

Table of Contents
Introduction: The New Frontier of Neuromodulation in Clinical Practice
Hello, and welcome to our educational platform. I’m Dr. Alex Jimenez, and I’m thrilled to share some transformative insights with you today. As a practitioner with credentials as a Doctor of Chiropractic (DC) and a Family Nurse Practitioner-Advanced Practice Registered Nurse (FNP-APRN), my career has been a journey dedicated to integrating diverse, evidence-based modalities to optimize patient outcomes. My passion is at the intersection of musculoskeletal health, neurology, and systemic wellness, expanding the possibilities in rehabilitative and restorative care. Today, we’re not just talking about treatment; we’re exploring the profound science of neuromodulation—the art and science of purposefully influencing the nervous system to achieve specific therapeutic goals. This field is rapidly evolving, driven by pioneering researchers who use sophisticated, modern methods to decode the language of our nerves. The findings are nothing short of revolutionary, offering us tools that are more precise, targeted, and powerful than ever before.
In this comprehensive educational post, I will present an evidence-based synthesis of how electricity interacts with neural tissue; why alternating current versus direct current matters for neuromodulation; how frequency, pulse width, and dose produce distinct clinical effects; and how these principles translate into everyday decision-making for pain relief, functional restoration, and neurorehabilitation. Drawing on the work of leading researchers in neuroplasticity, cortical-spinal synchronization, and paired associative stimulation, I aim to clarify misconceptions, highlight what is clinically safe and effective, and provide practical strategies you can apply in real-world settings. Across the past several decades, neuroscience has undergone a pivotal transformation. Early mechanistic theories—rooted in fluid-based conduction—yielded to the robust demonstration that the nervous system communicates via bioelectricity and ionic gradients. In the 1990s and early 2000s, converging experimental evidence established synaptic and network-level plasticity as a core organizing principle of brain function. Hebbian learning—summarized by the phrase “neurons that fire together, wire together”—was quantitatively explored through paradigms like paired associative stimulation, while stroke rehabilitation researchers advanced protocols that pair repetitive peripheral input with voluntary movement to strengthen pathways and drive recovery.
These discoveries invigorated clinical translation: peripheral and central neuromodulation (including vagus nerve stimulation and closed-loop wearable systems) now help retrain sensorimotor circuits, augment cortical output, and temper pathological signaling. Yet amidst this progress, technical misunderstandings persist—especially around the distinctions between alternating current and direct current, the risks of skin chemical shifts and faradic reactions, and the role of frequency relative to axonal refractory periods. We will begin by demystifying one of the most fundamental yet misunderstood concepts in electrotherapy: electrical impedance. We will explore why factors like skin hydration, subcutaneous fat, scar tissue, and even air gaps can dramatically alter the effectiveness of a treatment. I’ll explain Ohm’s Law in a clinically relevant way, illustrating how understanding resistance is crucial to delivering a safe and effective therapeutic dose of electricity to the target nerve or muscle without causing unnecessary pain. From there, we will explore the “levers of neuromodulation”—the specific parameters we can control to orchestrate a desired biological response. We’ll dissect the critical role of frequency. You will learn how different frequency bands can be used to achieve distinct clinical outcomes, from activating the body’s innate pain-relieving opioid pathways with low frequencies (long-term depression) to enhancing motor learning and neuroplasticity with mid-range frequencies (long-term potentiation). We will even touch upon how high frequencies are used for sensory gating in common TENS applications and how kilohertz frequencies can create outright nerve conduction blocks, a principle used in advanced procedures like radiofrequency ablation. A key concept we’ll unpack is neural entrainment, where we can synchronize brainwave activity by stimulating a peripheral nerve at a specific frequency, opening up incredible possibilities for improving everything from motor control and balance to cognitive focus and attention.
We will also investigate other critical parameters, such as pulse width and its role in selective nerve fiber activation—how we can target sensory fibers versus motor fibers—and amplitude (intensity), understanding the sequential recruitment of nerve fibers from sensory to motor to pain. This knowledge allows us to tailor our treatments with surgical precision, moving beyond a one-size-fits-all approach. Finally, we will cover the essential, non-negotiable safety protocols. I’ll outline the absolute and relative contraindications for peripheral nerve stimulation, emphasizing the critical importance of patient screening for conditions like pacemakers, deep brain stimulators, and pregnancy. We will discuss why certain anatomical areas, like the anterior neck over the carotid sinus, are “no-go zones” and the physiological reasons behind these precautions. This post is designed to be a thorough, narrative-based exploration of these concepts, providing you with the “why” behind the “what,” grounded in the latest evidence-based research. Let’s begin this journey into the electrifying world of neuromodulation.
Decoding Electrical Impedance: The First Hurdle in Effective Neuromodulation
When we use any form of electrotherapy, our primary goal is to deliver a controlled, predictable amount of electrical stimulation to a target tissue—be it a peripheral nerve, a muscle, or a specific sensory pathway. However, there’s a significant variable that stands between our device and that target: electrical impedance. Think of impedance as the total opposition a biological circuit presents to the flow of electrical current. It’s a more comprehensive term than simple resistance because it includes factors relevant to alternating currents, which are common in therapeutic devices.
 To truly grasp its importance, we need to revisit a fundamental principle of physics: Ohm’s Law. The formula is elegant in its simplicity: Voltage = Current × Resistance (or Impedance). In the context of our therapeutic devices, the voltage is often a fixed or limited output of the machine. The current, measured in amperage, is what actually creates the physiological effect—it’s the flow of electrons that depolarizes the nerve. This means that if the voltage from our device is relatively constant, the current that successfully reaches the nerve is inversely proportional to the impedance of the tissues it must cross.
To truly grasp its importance, we need to revisit a fundamental principle of physics: Ohm’s Law. The formula is elegant in its simplicity: Voltage = Current × Resistance (or Impedance). In the context of our therapeutic devices, the voltage is often a fixed or limited output of the machine. The current, measured in amperage, is what actually creates the physiological effect—it’s the flow of electrons that depolarizes the nerve. This means that if the voltage from our device is relatively constant, the current that successfully reaches the nerve is inversely proportional to the impedance of the tissues it must cross.
Let me put this information into a clinical scenario. The dry, outer layer of the skin, the stratum corneum, is an incredibly effective insulator. It can present an impedance of anywhere from 40,000 to 1,000,000 ohms. Let’s use Ohm’s Law again. If your impedance (R) is 100,000 ohms and your device’s voltage (V) is fixed, the current (I) that flows is incredibly small. To achieve a therapeutic effect at the target nerve, you would need to significantly increase the device’s current output. If the impedance is 100,000 times higher than ideal, you would also have to massively increase the amperage required. What happens when you turn the amperage up that high on a patient? They experience a sharp, unpleasant, and often painful sensation. Ouch! This is not a therapeutic sensation; it’s a nociceptive one, stimulating pain fibers and causing the patient to guard and resist the treatment.
This is precisely why managing impedance is not just a technical detail—it’s a cornerstone of effective and comfortable neuromodulation. So, how do we control it?
Practical Strategies for Overcoming High Impedance
The good news is that we have several effective methods to decrease impedance and ensure our therapeutic current reaches its intended destination. Understanding these variables is what separates a novice from an expert practitioner.
The Power of Hydration and Conductive Gels
The single most effective way to reduce skin impedance is to wet the skin. Simply applying water can drop impedance by a factor of 10 to 100. This is because water, especially when it contains electrolytes, is an excellent conductor of electricity. However, water evaporates. This is where conductive gels and solutions come in. The primary purpose of a lubricant or gel is not just to help the electrode glide over the skin; its most crucial role is to fill in the microscopic air gaps between the electrode and the skin.
 Electricity does not travel well through air. Air is a fantastic insulator. When electricity tries to cross an air gap, it needs to accumulate a massive electrical potential before it can discharge across that gap. What do we call a massive, uncontrolled discharge of electricity through the air? Lightning. Now, imagine tiny lightning strikes happening on your patient’s skin between the electrode and their epidermis. This creates a sharp, stinging, and erratic sensation. Gels eliminate these air gaps, creating a continuous, low-impedance pathway for the current to flow smoothly and comfortably into the tissues. Furthermore, factors like skin oils and fine hairs also contribute to impedance, and a good gel helps to bridge these as well. With well-prepped electrodes and proper skin preparation using a conductive medium, we can bring the impedance down from tens of thousands of ohms to as low as 500 ohms. This is a monumental difference and is key to a successful treatment.
Electricity does not travel well through air. Air is a fantastic insulator. When electricity tries to cross an air gap, it needs to accumulate a massive electrical potential before it can discharge across that gap. What do we call a massive, uncontrolled discharge of electricity through the air? Lightning. Now, imagine tiny lightning strikes happening on your patient’s skin between the electrode and their epidermis. This creates a sharp, stinging, and erratic sensation. Gels eliminate these air gaps, creating a continuous, low-impedance pathway for the current to flow smoothly and comfortably into the tissues. Furthermore, factors like skin oils and fine hairs also contribute to impedance, and a good gel helps to bridge these as well. With well-prepped electrodes and proper skin preparation using a conductive medium, we can bring the impedance down from tens of thousands of ohms to as low as 500 ohms. This is a monumental difference and is key to a successful treatment.
The Influence of Tissue Composition on Current Flow
Once the current bypasses the skin, its journey is further influenced by the composition of the underlying tissues. Different tissues have different conductive properties, largely based on their water and electrolyte content.
- Subcutaneous Fat (Adipose Tissue): Fat is a poor conductor of electricity and has a significantly higher impedance than muscle. This has direct clinical implications. If I am treating a patient with a higher degree of adiposity, particularly over the treatment area like the legs or lower back, I know I will need to increase the intensity (amplitude) on my device to get the same level of nerve or muscle activation. You’ve likely observed this in your own practice. When you’re trying to achieve a motor contraction on a patient with very lean, muscular legs, a small amount of current can make them nearly twitch off the table. Conversely, for a patient with a thick subcutaneous fat layer, you might find yourself needing to “crank the dial up” considerably to see any response.
- Muscle Tissue: In contrast, muscles are excellent conductors. They are rich in water and electrolytes (like sodium, potassium, and calcium), which are essential for conducting the electrical signals that cause contraction. This makes them relatively easy targets for stimulation once you get past the overlying skin and fat.
- Bone: Bone is a very poor conductor. It is dense and has low water content, presenting a high-impedance barrier to current flow. This is important to consider when placing electrodes, as current will preferentially travel around bone rather than through it.
- Scar Tissue: This is a critically important one for us in rehabilitative medicine. Scar tissue is a notoriously bad conductor. It’s dense, fibrotic, and lacks the organized, water-rich structure of healthy tissue. When you are treating an area with significant scarring, perhaps from a previous surgery or injury, you must anticipate a much higher impedance. This can make it very difficult to stimulate nerves or muscles underneath or within that scar. Sometimes, the therapeutic goal might even be to work on the scar tissue itself, but we must be aware that it will dramatically alter the path and intensity of the electrical current.
This concept of bypassing resistance is precisely why modalities like needle acupuncture or electromyography (EMG) with needle electrodes are so effective at eliciting a response. By inserting a fine needle, they completely bypass the high-impedance barrier of the skin, delivering the electrical current directly to the target nerve or muscle. This allows for an incredibly efficient and precise stimulation with very low levels of current.
Neurophysiology Foundations: From Fluids to Bioelectricity
I grew up, clinically speaking, in a world where certain ancient ideas lingered—ideas proposing that nerves conducted “fluids” rather than electricity. The maturation of electrophysiology dismantled those notions. Today, we recognize that neurons are exquisitely tuned electrochemical machines: ion channels set resting membrane potentials, spikes emerge when depolarization crosses threshold, and synaptic plasticity reshapes circuits in response to repeated patterns of activation.
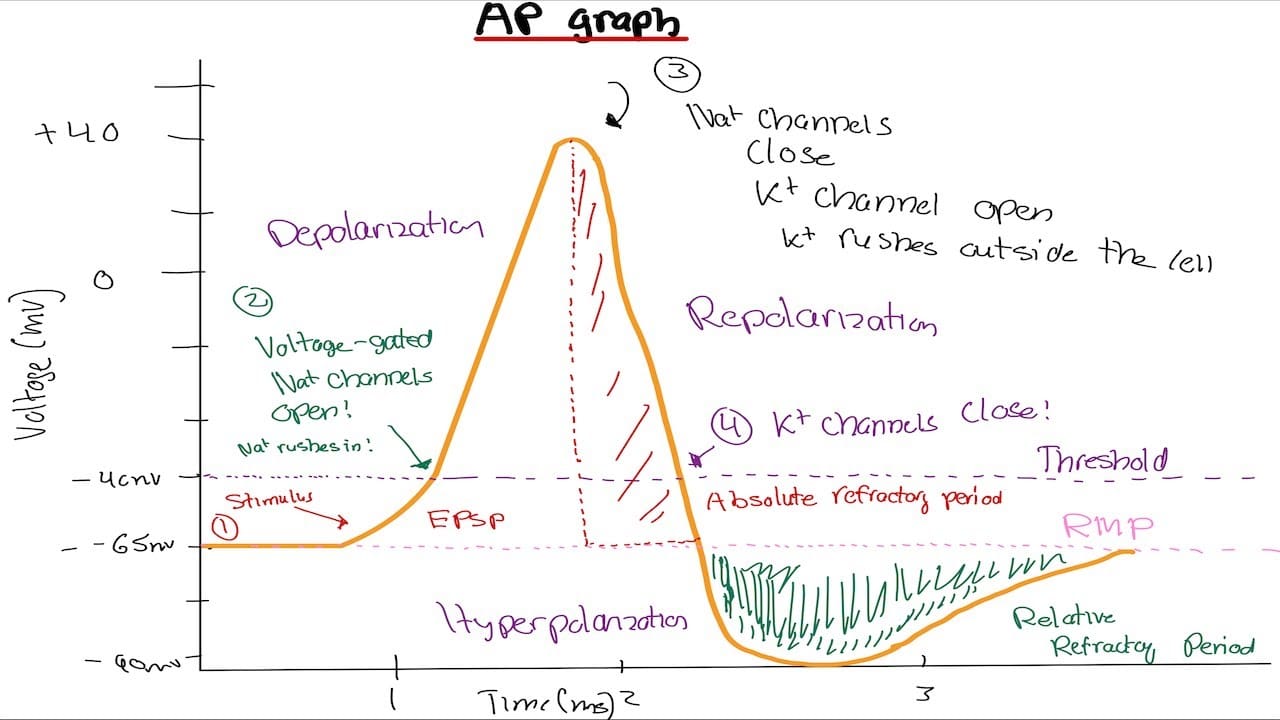
At the cellular level:
- The resting membrane potential (~-70 mV in many neurons) arises from selective permeability (especially to potassium) and active maintenance by sodium-potassium ATPase.
- Voltage-gated sodium channels open during depolarization, allowing Na+ influx, driving the rising phase of the action potential.
- Voltage-gated potassium channels open later, facilitating K⁺ efflux that repolarizes the membrane and eventually hyperpolarizes it.
- Refractory periods follow each spike: an absolute refractory period (about 1–2 ms) during which no stimulus can trigger another spike, and a relative refractory period (about 2–5 ms) during which a stronger-than-usual stimulus is needed.
These timing properties bound the maximum firing rates of axons (often between 500 and 1000 Hz) and become critical when we choose stimulation frequencies. If we drive a neuron during its refractory period, we impede conduction; if we time pulses to allow recovery, we facilitate firing and enable plastic changes that accumulate with repetition.
Hebbian Learning and Paired Associative Stimulation
Modern neuroplasticity research showed that when two neurons repeatedly fire together, synaptic efficacy between them strengthens—“fire together, wire together.” In humans, paired associative stimulation (PAS) operationalizes this by delivering temporally coordinated inputs—for example, a peripheral nerve stimulus paired with transcranial stimulation or voluntary movement. When the timing aligns so that the cortical and spinal volleys converge within an effective window, synapses and pathways potentiate.
 Clinically, this matters profoundly:
Clinically, this matters profoundly:
- In stroke rehabilitation, pairing peripheral stimulation of the affected limb with task-specific voluntary movement enhances corticospinal synchronization and improves motor output.
- Closed-loop wearable systems detect movement or physiological signals and deliver stimulation in synchrony, amplifying relevant activity while minimizing noise.
- The principle is repetition with timing: repeated volleys produce durable changes that we enlist to rebuild function.
The Levers of Neuromodulation: Tailoring Treatment with Frequency, Pulse Width, and Amplitude
Once we have managed impedance and established a clear path for our current, we can begin to manipulate the characteristics of the electrical signal itself to achieve specific therapeutic goals. Think of these parameters as the “levers” or “dials” of neuromodulation. The three most fundamental levers are frequency, pulse width, and amplitude. More is not always better; different is better.
 Frequency: The Key to Therapeutic Intent
Frequency: The Key to Therapeutic Intent
Frequency, measured in Hertz (Hz), refers to the number of pulses or cycles of electricity delivered per second. This is arguably the most important parameter for determining the type of physiological response you will create. Different frequencies preferentially activate different neural pathways and mechanisms. Based on decades of research from leading neurophysiologists, we can establish some general rules of thumb:
 Low Frequency (1-10 Hz): Stimulation in this range has been consistently shown to activate the body’s endogenous opioid pathways. When we stimulate sensory nerves at these low frequencies, we trigger the release of endorphins and enkephalins in the brainstem and spinal cord. This doesn’t just block pain signals; it actively suppresses the pain pathways. The technical term for this sustained calming effect on neuronal activity is Long-Term Depression (LTD). It’s important to clarify that this is not psychological depression; in neurophysiology, LTD refers to a long-lasting reduction in the strength of synaptic connections. We are essentially teaching overactive, sensitized pain circuits to calm down by promoting protein synthesis in neurons, which reduces excitability. This is a healthy, active process, distinct from a simple nerve block.
Low Frequency (1-10 Hz): Stimulation in this range has been consistently shown to activate the body’s endogenous opioid pathways. When we stimulate sensory nerves at these low frequencies, we trigger the release of endorphins and enkephalins in the brainstem and spinal cord. This doesn’t just block pain signals; it actively suppresses the pain pathways. The technical term for this sustained calming effect on neuronal activity is Long-Term Depression (LTD). It’s important to clarify that this is not psychological depression; in neurophysiology, LTD refers to a long-lasting reduction in the strength of synaptic connections. We are essentially teaching overactive, sensitized pain circuits to calm down by promoting protein synthesis in neurons, which reduces excitability. This is a healthy, active process, distinct from a simple nerve block.- Medium Frequency (20-50 Hz): This range is the sweet spot for motor applications and inducing neuroplasticity. When we stimulate at these frequencies, we can achieve strong, tetanic muscle contractions for strengthening (motor recruitment). More excitingly, this frequency band, particularly around 40 Hz (gamma band), is synonymous with inducing Long-Term Potentiation (LTP). LTP is the cellular mechanism underlying learning and memory. It involves strengthening the synaptic connections between neurons. In a rehabilitation setting, this means we can use 40 Hz stimulation to enhance motor learning, improve motor timing, and “rewire” the brain to create more efficient and stable movement patterns.
- High Frequency (80-150 Hz): This is the range most commonly used by conventional Transcutaneous Electrical Nerve Stimulation (TENS) units. The mechanism here is primarily sensory gating and analgesia, based on the Gate Control Theory of Pain proposed by Melzack and Wall. By stimulating the large, fast-conducting A-beta sensory fibers, we effectively “close the gate” in the spinal cord, preventing the signals from the smaller, slower pain fibers (A-delta and C-fibers) from reaching the brain. This provides rapid, but often temporary, pain relief.
- Kilohertz Frequency (kHz): When we get into the range of thousands of hertz, the mechanism changes again. These ultra-high frequencies can create a true nerve conduction block. The nerve membrane becomes unable to repolarize quickly enough between pulses, leading to a complete, albeit temporary, shutdown of signal transmission. This is the principle behind certain advanced pain management techniques like radiofrequency ablation, where a targeted nerve is essentially silenced.
The key takeaway here is that frequency determines the therapeutic intent. You must first decide what your clinical goal is—opioid-mediated pain relief, motor learning, or sensory gating—and then select the appropriate frequency.
Pulse Width and Amplitude: Refining Your Target
While frequency sets the overall therapeutic goal, pulse width and amplitude allow us to refine our target and control the intensity of the stimulation.
Pulse Width (Pulse Duration): The Key to Fiber Selection
Pulse width, measured in microseconds (µs), refers to the duration of each individual electrical pulse. This parameter is crucial for achieving selective activation of different nerve fiber types. Different nerves have different “chronaxies,” or minimum pulse durations required to excite them.
- Short Pulse Width (e.g., 50-100 µs): Shorter pulse durations tend to be more comfortable and will preferentially activate the large, myelinated A-beta sensory fibers. This is ideal if your goal is purely sensory, such as for sensory gating (TENS) or providing non-painful feedback to the nervous system.
- Longer Pulse Width (e.g., 200-400 µs): As you widen the pulse, you start to more effectively recruit the A-alpha motor fibers. This is what you need to elicit a muscle contraction. If your goal is muscle strengthening, re-education, or inducing LTP through motor activation, you’ll need a wider pulse width.
- Very Long Pulse Width (e.g., >400 µs): At even longer pulse durations, you begin to activate the smaller, higher-threshold A-delta and C pain fibers. This will start to feel sharp and painful.
Some devices allow you to set the pulse width, while others have it fixed. If you have the ability to control it, it gives you another powerful lever to ensure you are stimulating the right nerve fibers for your specific clinical goal.
Amplitude (Intensity): The Master Control
Amplitude refers to the intensity of the current, typically controlled by the “intensity” dial on the device. While pulse width helps with selectivity, you can always override this selectivity by increasing the amplitude. The recruitment of nerve fibers almost always occurs in a sequential, predictable order as you turn up the intensity:
- Sensory (A-beta fibers): The first thing the patient will feel is a gentle tingling or buzzing sensation. This is the activation of the large sensory fibers.
- Motor (A-alpha fibers): As you increase the intensity further, you will reach the motor threshold, and you will begin to see a visible muscle twitch or contraction.
- Pain (A-delta and C-fibers): If you continue to increase the intensity, you will eventually activate the nociceptive (pain) fibers, and the sensation will become unpleasant, sharp, or painful. Any device, if turned up high enough, will eventually cause pain.
So, the crucial question becomes: Do we ever want to stimulate pain fibers? The answer, surprisingly, is not always “no.” Pain is a very powerful limbic and emotional motivator. It creates a strong state of arousal. Consider a patient with a condition like narcolepsy, who has difficulty staying awake. If you apply a gentle sensory current, it might actually be soothing and help them fall asleep faster. A motor-level current might not be enough to keep them aroused. But a brief, sharp, painful stimulus? That will certainly wake them up and increase their level of alertness.
However, in the vast majority of our cases, especially with patients suffering from chronic neuropathic pain, the last thing we want to do is stimulate more pain fibers. For these individuals, our goal is to provide relief and calm the nervous system, not to further sensitize it. For neuromodulation aimed at creating plasticity and motor learning, we typically want to be at a strong sensory or comfortable motor level, well below the pain threshold.
Alternating Current vs. Direct Current: Mechanisms and Clinical Implications
 Understanding AC versus DC is non-negotiable for safe, effective neuromodulation.
Understanding AC versus DC is non-negotiable for safe, effective neuromodulation.
- Alternating Current (AC):
- The polarity reverses periodically; the anode and cathode swap roles over time.
- Modern AC neurostimulation employs charge-balanced, biphasic, symmetrical waveforms. Because polarity alternates, net charge over time tends toward zero at the skin interface.
- Clinical impact: Minimal faradic reactions and minimal pH shifts in the skin reduce the risk of irritation or burns. AC is excellent for dynamic neuromodulation—sending repeated, timed volleys that facilitate plasticity while maintaining tissue safety.
- Direct Current (DC):
- Constant polarity (current flows from anode to cathode).
- Sustained DC can create steady ionic displacement, leading to electrochemical gradients and capacitive effects in tissues. This property makes DC useful for iontophoresis (drug delivery) and certain specialized central applications like transcranial direct current stimulation (tDCS) at very low amplitudes over long durations.
- Risks peripherally: DC at clinically meaningful amplitudes over peripheral nerves can increase local pH shifts, gas formation, and skin irritation; it can also block or ablate nerve conduction at higher frequencies or settings (as with radiofrequency neurotomy). Therefore, DC is generally not appropriate for peripheral neuromodulation aimed at facilitating neuroplasticity.
An analogy helps: a river carving a canyon reflects the erosive potential of persistent unidirectional flow (DC), whereas waves on a beach reflect bidirectional movement with far less net erosion (AC). In neuromodulation, we prefer the “beach” scenario—safe, oscillatory delivery that avoids charge accumulation and chemical damage.
How Electric Stimulation Actually Couples to Tissue
A common misconception is that stimulation “pumps electrons” into the body. In reality:
- Electrons congregate on the electrode surface, not in the body.
- The interface requires a conductive medium (e.g., saline, ultrasound gel) containing ions such as Na⁺, Cl⁻, K⁺.
- When a potential is applied, ions in the medium align: positive ions are attracted toward negatively charged electrode surfaces and vice versa.
- This ionic organization establishes an electric field (E-field) across tissue layers: epidermis, dermis, subcutaneous tissue, muscle, vessels, and nerves.
Within neurons:
- The E-field influences voltage-gated sodium channels through their S4 voltage-sensor domains, which respond to changes in transmembrane electric potential.
- Depolarization occurs when Na+ influx increases, raising the membrane potential toward threshold. If sufficient depolarization occurs (e.g., shifting from ~-70 mV toward 0 mV), an action potential fires and propagates.
This is why conductivity matters:
- Poor gel application, dehydrated skin, or electrolyte imbalance can degrade coupling and reduce clinical effect.
- Overhydration or deranged electrolytes can also distort ionic behavior, sometimes rendering stimulation ineffective or uncomfortable. In clinical practice, we assess hydration and overall health status, especially in medically complex patients, before and during neuromodulation.
Refractory Periods and Frequency: Facilitating vs. Blocking
The timing structure of action potentials sets the rules for frequency selection:
- Absolute refractory period (~1–2 ms): no stimulus can re-fire the neuron.
- Relative refractory period (~2–5 ms): a stronger stimulus than baseline can re-fire the neuron.
From these intervals, typical maximum axonal firing rates span ~500–1000 Hz. Clinically:
- Very high frequencies (approaching or exceeding refractory overlap) tend to impede conduction and can block signaling. Radiofrequency neurotomy leverages this to ablate painful pathways; it is not used to facilitate cortical-spinal plasticity.
- Low to mid frequencies (≈1–100 Hz) align with axonal recovery timing and are optimal for plastic facilitation—repeated, recoverable volleys that strengthen pathways through Hebbian mechanisms.
- Pain management can use gating strategies, often in low and mid ranges, to modulate dorsal horn processing and thalamocortical relay of nociception.
Thus, the clinical intent dictates frequency:
- To reduce neuropathic pain via block: higher frequencies (medical procedures, not routine peripheral neuromodulation).
- To facilitate movement, learning, and recovery: low- to mid-frequency AC stimulation synchronized with voluntary tasks.
Parameter Mastery: Voltage, Current, Impedance, Pulse Width, Frequency, and Charge
Clinically, we must translate physics into therapeutics.
- Voltage (V): akin to pressure—helps overcome skin impedance and allows the field to establish across tissue.
- Current (I) or Amperage (mA): determines recruitment, i.e., how many axons are engaged.
- Impedance (R): opposition to current flow—affected by skin properties, hair, callus, hydration, and whether you use conductive gel or saline.
- Pulse Width (PW): duration of each pulse—longer widths deliver more charge per pulse, often recruiting deeper or larger fibers, but may increase discomfort.
- Frequency (Hz): pulses per second—governs firing patterns: low/mid for plastic facilitation; higher for gating or block.
- Charge (Q): Q = Current × Pulse Width—this is the true dose per pulse. Therapists often overlook charge, but it directly reflects how much energy we deliver to tissue.
Ohm’s Law (V = I × R) guides expectations:
- On most clinical devices, voltage is fixed or semi-fixed; resistance is dictated by the patient and setup; current is our adjustable control.
- Improve coupling (reduce R) with good skin prep and conductive media; then titrate current to desired recruitment while respecting comfort and safety.
- Adjust pulse width to fine-tune recruitment depth and perception.
- Choose frequency based on the clinical goal (facilitation vs. gating vs. block).
Skin Safety, pH Shifts, and Faradic Reactions
Chemistry at the skin-electrode interface is as important as physics:
- DC can cause net ionic drift, producing acid-base imbalances and faradic reactions. Prolonged DC exposure may lead to pH shifts, gas formation, and skin irritation or burns.
- AC with charge-balanced waveforms minimizes net charge accumulation, thereby reducing chemical damage and improving tolerability.
Clinical pearls:
- Use proper gel or saline.
- Avoid prolonged DC application over peripheral nerves when the goal is neuromodulation for plastic facilitation.
- Monitor skin before, during, and after sessions; rotate electrode sites when appropriate.
- Keep electrode surfaces clean and intact; worn electrodes can produce hotspots and uneven current density.
Device Selection: Determining AC vs. DC and Reading Waveforms
To determine whether a device is AC or DC:
- Check the device label for “Output: AC” or “Output: DC.”
- Review the manual for keywords:
- AC indicators: balanced charge, biphasic, symmetrical waveform.
- DC indicators: galvanic, monophasic, iontophoresis.
- Inspect waveform graphs:
- DC traces do not cross zero (constant polarity).
- AC traces alternate, crossing the zero line regularly.
If your clinical aim is peripheral neuromodulation to facilitate plasticity, choose AC with charge-balanced biphasic waveforms and appropriate low to mid frequencies.
Peripheral vs. Central Stimulation: Where DC Fits
While DC is not ideal for peripheral neuromodulation aimed at facilitation, it has specific roles:
- Transcranial Direct Current Stimulation (tDCS): very low amplitude DC applied over long durations can modulate cortical excitability through subtle shifts in membrane potential. This is a central application with carefully controlled parameters and safety protocols.
- Nerve blocks and radiofrequency ablation: DC-like or high-frequency radiofrequency procedures aim to inhibit or ablate painful pathways. These are interventional medical procedures, not plastic-facilitation therapies.
For peripheral facilitation—wrist extensors, median nerve inputs, vagus nerve augmentation to support autonomic balance—AC with appropriate frequencies is the safer, more effective choice.
Clinical Translation: Pairing Peripheral Stimulation with Voluntary Movement
Pairing alternating current peripheral stimulation with voluntary movement amplifies relevant neural signals:
- Apply a nerve stimulator at the wrist while the patient performs task-specific movements of the hand and forearm.
- Time pulses to occur in proximity to movement initiation or the phase of maximal intended activation. Repeated pairing strengthens corticospinal pathways via Hebbian mechanisms.
- In vagus nerve stimulation paradigms, use low- to mid-frequencies that do not push the nerve into block; coordinate with breathing or motor tasks to maximize autonomic and sensorimotor benefits.
Over weeks, these protocols improve output—grip strength, dexterity, and coordination—and enhance sensory integration, especially after a stroke or neuropathic injury.
Pain Modulation: Gating vs. Blocking
Pain strategies differ:
- Gating (e.g., conventional TENS): uses frequencies and pulse widths that activate non-nociceptive Aβ fibers, inhibiting nociceptive transmission in the dorsal horn via interneuronal circuits. Typically low to mid frequencies with modest amplitudes aligned to comfort.
- Blocking/ablation: interventional procedures at high frequencies to disrupt pathological firing; not used to facilitate plasticity.
Always align the intended outcome with the frequency band and waveform:
- For temporary relief and functional enablement: gating with AC.
- For long-term facilitation: AC with low/mid frequency pairing and task-specific training.
- For refractory pain requiring procedural intervention: medical block/ablation.
Neural Entrainment: Synchronizing Brainwaves with Peripheral Stimulation
One of the most exciting frontiers in neuromodulation is a concept called neural entrainment. This is where we can actually influence and synchronize the brain’s own rhythmic electrical activity—its brainwaves—by delivering a rhythmic stimulus to a peripheral nerve. It’s a truly remarkable phenomenon. When we stimulate a peripheral nerve, say the median nerve at the wrist or the trigeminal nerve on the face, the electrical signals travel up the spinal cord to the thalamus, the brain’s central relay station. From the thalamus, these signals project out to the cerebral cortex and other brain regions.
If we deliver this peripheral stimulation at a consistent, rhythmic frequency that matches one of the brain’s natural resonant frequencies, the brain’s neuronal populations will start to fire in sync with our external stimulus. This thalamocortical connection allows us to “tune” the brain to a desired state. Let’s look at some examples:
- Theta Band (4-8 Hz): The theta rhythm is strongly associated with the cerebellum, a key area for motor learning, error correction, and sensory-motor calibration. If I have a patient with poor motor patterns—perhaps they have an unstable gait, poor posture, or general clumsiness—I can deliver peripheral nerve stimulation at a theta frequency (e.g., 6 Hz). This signal will entrain the cerebellum, helping to stabilize its function and improve balance and motor control. It’s like providing a metronome for the part of their brain that coordinates movement.
- Mu and Alpha Bands (8-12 Hz): The mu rhythm is specifically linked to the sensorimotor cortex and is involved in sensory-motor binding—the integration of sensory feedback with motor commands. The alpha rhythm is associated with sensory gating, cortical inhibition, and a state of “readiness.” It indicates a brain that is calm but alert, ready for the next incoming stimulus. Modulating in this range can help in conditions where there is a disconnect between sensory perception and motor output.
- Beta and Gamma Bands (20-40 Hz): As mentioned earlier, this is the realm of Long-Term Potentiation (LTP), motor timing, and higher cognitive functions like attention. Do you want to help a patient pay attention better? You can apply a nerve stimulator to their median nerve and set it to 40 Hz (gamma frequency). The rhythmic input will travel to their cortex and help synchronize the neurons involved in perceptual processing and attention. This can enhance their ability to focus and process information from their environment. It’s an incredible application of basic neuroscience.
This is the kind of targeted intervention that demonstrates the unique power of neuromodulation. These are effects we simply cannot achieve with a chiropractic adjustment, for example. We are not adjusting a joint at 40 times per second. This isn’t to say one is better than the other; it highlights how these tools are wonderfully complementary. We can use an adjustment to restore joint mechanics and then use neuromodulation to retrain the neural pathways that control that joint. It’s a truly integrated approach.
A Practical Framework for Setting Your Device
Let’s bring this all together into a simple, step-by-step clinical thought process for setting up a neuromodulation treatment.
- Determine Your Target Tissue: What are you trying to stimulate? Is it a superficial sensory nerve? A deep muscle? Are you trying to avoid a nerve and just work on a muscle belly? Your electrode placement and initial parameter thoughts will start here.
- Determine Your Therapeutic Goal: This is the most important step. What is the physiological outcome you want to achieve?
- Do you want to suppress pain via opioid release? (Low frequency, LTD).
- Do you want to activate a muscle and promote neuroplasticity? (Medium frequency, LTP).
- Do you want to provide temporary analgesia via sensory gating? (High frequency).
- Do you want to block a nerve completely? (Kilohertz frequency).
- Determine Your Treatment Time and Duty Cycle: How long will the treatment last? Will the stimulation be continuous, or will it be intermittent (e.g., 10 seconds on, 10 seconds off)? Duty cycles are particularly important for muscle strengthening to prevent fatigue.
- Choose Your Parameters and Intensity: Based on your goals, you will now set your frequency, pulse width (if available), and finally, your amplitude. You’ll slowly increase the intensity, communicating with the patient, until you reach the desired sensory or motor threshold that aligns with your therapeutic goal, always staying below the threshold of pain unless there is a specific and rare reason to exceed it.
Practical Steps: Setting Up a Safe, Effective Session
- Skin preparation:
- Clean the area; remove oils and lotions.
- Apply conductive gel or saline-soaked pads to lower impedance.
- Inspect skin integrity and avoid broken or irritated sites.
- Electrode placement:
- Position near the target nerve or muscle belly relevant to the clinical goal.
- Maintain firm, even contact; avoid hotspots by using appropriately sized electrodes.
- Parameter selection:
- Waveform: AC, charge-balanced biphasic.
- Frequency: start in 1–20 Hz for facilitation and adjust up to 50–100 Hz based on tolerance and goals; avoid very high frequencies for peripheral facilitation.
- Pulse width: moderate values to balance recruitment and comfort.
- Current: titrate to a strong-but-comfortable perception; observe for visible, non-painful muscle activation if desired.
- Task pairing:
- Coordinate stimulation with specific movements or task practice for 10–30 minutes depending on tolerance and program design.
- Use rest intervals to prevent fatigue, especially in neurologic populations.
- Monitoring and documentation:
- Record baseline and post-session outcomes.
- Adjust parameters over sessions based on response, comfort, and functional gains.
Troubleshooting and Optimization
- If stimulation is uncomfortable or ineffective:
- Recheck conductive medium and electrode contact.
- Verify device output type (AC vs. DC) and waveform (charge-balanced biphasic).
- Adjust pulse width and current downward if discomfort persists; increase gradually once tolerated.
- Ensure frequency is in the low to mid range; avoid refractory overlap.
- Confirm the patient’s hydration and electrolyte status; consider medical assessment if symptoms suggest dysregulation.
- If you suspect blocking instead of facilitation:
- Reduce frequency.
- Shorten pulse width or reduce current.
- Reassess timing relative to voluntary movement.
- If you cannot determine device output:
- Contact the manufacturer.
- Review technical documentation.
- Prefer devices with transparent specifications and clinical support.
Critical Safety: Contraindications for Peripheral Nerve Stimulation
As with any powerful modality, the responsible use of peripheral nerve stimulation requires a thorough understanding of its contraindications. Patient safety must always be our highest priority. We can categorize these contraindications into absolute (never do it) and relative (proceed with extreme caution and clinical judgment).
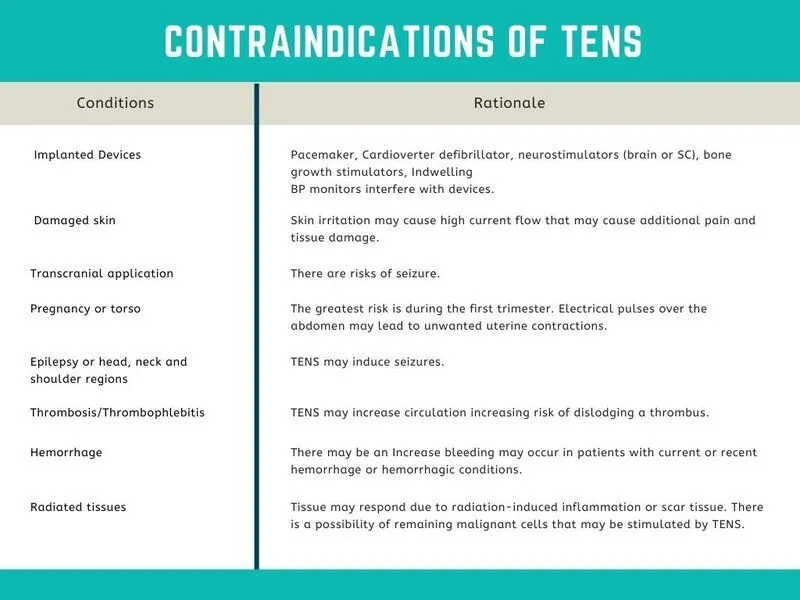 Absolute Contraindications
Absolute Contraindications
These are situations where peripheral nerve stimulation should never be used:
- Over or Near Implanted Electronic Devices: This includes pacemakers, implantable cardioverter-defibrillators (ICDs), cochlear implants, and deep brain stimulators (DBS). The electrical current from your device can interfere with the function of these life-sustaining devices, with potentially fatal consequences.
- Over the Anterior Neck / Carotid Sinus: This is a critical “no-go zone.” The carotid sinus contains baroreceptors that provide direct input to the brain about blood pressure. Stimulating this area can trick the brain into thinking blood pressure is dangerously high, triggering a powerful reflex that can cause pathological bradycardia (a dangerously slow heart rate) or even an atrioventricular (AV) block, where the heart’s electrical conduction system is disrupted. This can cause the heart to seize up. It is simply not worth the risk. It’s interesting to note that some newer forms of vagus nerve stimulation are moving away from the neck and instead targeting the auricular branch of the vagus nerve in the ear. This provides input to the vagus nucleus in the brainstem without the risk of direct cardiac interference.
- Directly Over Reproductive Organs: This includes areas like the scrotum, vulva, and testes. The effects on these sensitive tissues and gametes are not well-studied and should be avoided.
- Over Open Wounds, Broken Skin, or Active Infections: Applying electrical current to these areas can worsen the condition, impede healing, and potentially spread infection.
- Near or Over Areas of Known or Suspected Malignancy (Cancer): The concern is that electrical stimulation could increase metabolic activity and blood flow in the tumor, potentially promoting its growth or metastasis. This remains a topic of research, but the standing recommendation is to avoid it.
- Over Regions of Deep Vein Thrombosis (DVT): Applying electrical current, especially if it causes muscle contraction, could dislodge a blood clot, leading to a life-threatening pulmonary embolism.
Relative Contraindications
These are situations that require careful consideration and often consultation with the patient’s primary medical team. The risks and benefits must be carefully weighed.
- Pregnancy: This is a major area of caution. While many people have found relief from low back pain during pregnancy using TENS, the general recommendation is to avoid abdominal, lumbar, and pelvic placements. I personally am extremely cautious when treating pregnant individuals. The stakes are incredibly high. Even if your intervention is completely unrelated to a subsequent complication, like a miscarriage, in the patient’s mind and potentially in a legal context, you may be held responsible. It’s a risk that I am not willing to take. A close friend of mine went through seven rounds of IVF and finally gave up. When we consider the emotional and physical journey that many people go through to conceive, we must be exceptionally mindful of any intervention, no matter how benign it may seem.
- Cognitive Impairment: If a patient cannot give you reliable feedback about what they are feeling, it is very difficult to safely dose the stimulation.
- Epilepsy: While some forms of neuromodulation are used to treat epilepsy, indiscriminate stimulation could potentially lower the seizure threshold in susceptible individuals.
Evidence-Based Safety and Contraindications in Peripheral and Electrical Neuromodulation
In my clinical practice, safety is the first principle. Before introducing any electrical nerve stimulation or peripheral neuromodulation, I systematically evaluate for absolute and relative contraindications. The goal is to prevent harm and preserve patient autonomy while leveraging proven benefits of neurostimulation.
- Absolute contraindications (context-dependent):
- Epilepsy: I avoid unmonitored stimulation in individuals with a seizure history unless under strict medical oversight and established protocols. Electrical inputs can alter cortical excitability, and frequency and intensity can impact seizure thresholds.
- Cardiac disease: Pacemakers, implantable cardioverter-defibrillators (ICDs), and unstable arrhythmias require cardiology involvement. Electromagnetic interference and autonomic shifts can be problematic. For non-implanted cardiac conditions, I monitor heart rate, blood pressure, and symptoms closely, and I start with low intensities and conservative sites away from the chest.
- Severe cognitive impairment or inability to report discomfort: When patients cannot communicate pain or adverse sensations (e.g., post-hypoxic injury after near-fatal drowning), stimulation requires enhanced caution. I often apply identical parameters to myself first to sense tolerability before any application, and I use minimal intensities with continuous observation.
- Relative contraindications:
- Neuropathy and regions of sensory loss: Stimulation may be less perceived or poorly localized. While not an absolute barrier, I adjust amplitudes, use alternative sites, and rely on objective autonomic measures (capillary refill and skin temperature asymmetries) rather than subjective sensation alone.
- Metallic implants or orthopedic hardware: Potential local heating, current shunting, or altered impedance are considerations. I avoid placing electrodes over hardware; I route stimulation through safer, contralateral or proximal sites and employ conservative duty cycles.
- Recent fractures and acutely inflamed tissues: I avoid direct stimulation over areas with edema, hematoma, or unstable injuries. When modulation is necessary for pain, I use remote sites and lower frequencies with short durations.
- Concurrent electrical medical devices: I avoid stacking unknown inputs. I coordinate with the patient’s care team to ensure device compatibility.
- Pregnancy: While hydrogen supplementation anecdotes circulate, I refrain from claims and avoid non-essential electrical interventions. If any supportive modality is considered, it must be cleared by obstetric providers, with risk-benefit clearly documented.
Why this cautious approach? Neuromodulation can alter autonomic balance, cortical excitability, and vascular dynamics. A careful intake, baseline vitals, and mapping of risks allow evidence-based, patient-specific decisions. This safety-first stance aligns with modern protocols in neuromodulation research and clinical practice.
Labeled Line Theory and Its Clinical Relevance in Peripheral Nerve Stimulation
The labeled line theory—originating in 19th-century sensory physiology—remains foundational. A labeled line is a dedicated pathway: stimulate a specific peripheral nerve, and you propagate signals along its characteristic central relay, activating defined nuclei and cortical targets.
- Why it matters clinically: When I stimulate the trigeminal nerve distribution, I’m not merely affecting local sensation. I’m engaging the trigeminal nucleus, relaying through the thalamus to the sensory cortex, and sending collaterals to the reticular formation. That collateralization heightens attention, arousal, and cortical excitability—useful in lethargy, attentional deficits, or post-concussive states.
- Median nerve stimulation is one of the most investigated PNS modalities. Afferent volleys ascend to the dorsal columns, thalamus, and sensorimotor cortex, producing robust cortical activation and motor priming. This labeled line is valuable across conditions: Tourette’s syndrome, concussion, Parkinson’s disease, and Alzheimer’s disease—where timing, activation, and network binding are therapeutic targets.
- Tibial nerve stimulation engages spinal afferents with downstream modulation of pain circuits (including endogenous opioids) and bladder control pathways. Evidence is substantial in overactive bladder and incontinence, demonstrating how distal stimulation can recalibrate central autonomic networks.
- Auricular vagus nerve stimulation (aVNS) follows a labeled line through the trigeminal nucleus, nucleus tractus solitarius, locus coeruleus, insula, and anterior cingulate cortex—regions central to parasympathetic tone, inflammation regulation, and affective-autonomic integration. By targeting the auricular branch, we can non-invasively modulate vagal circuitry.
Reasoning: Stimulating a labeled line is akin to dialing a direct number into a network hub. We choose a nerve based on its proven central relays and clinical goals—whether enhancing attention (trigeminal collaterals), priming the motor cortex (median), modulating bladder networks (tibial), or boosting parasympathetic coherence (auricular vagus).
Peripheral Stimulation as Brain Treatment: Autonomic, Limbic, and Cortical Modulation
A common misconception is that peripheral stimulation treats only the periphery. In reality, every felt stimulus is a central event. If you perceive the stimulation, I’ve altered your brain—a change in EEG coherence, autonomic output, or blood flow can occur even when sensations are subtle.
- Physiologic basis:
- Ascendant afferents via spinal and cranial nerve pathways increase thalamocortical resonance—the rhythmic coupling between thalamic pacemakers and cortical columns.
- Brainstem relays engage limbic circuits (amygdala, hippocampal formations) through NTS and LC-mediated norepinephrine modulation, influencing vigilance, stress reactivity, and pain perception.
- Frequency-dependent plasticity: Low frequencies (around 1–10 Hz) can drive long-term depression (LTD) and analgesia via endogenous opioids; mid-range frequencies (20–40 Hz) enhance sensory-motor binding, alertness, and short-term potentiation of cortical excitability.
- Durability: Immediate changes signify state shifts—valuable for priming rehabilitation or resetting maladaptive loops. To enhance persistence, I pair stimulation with task-specific movement, breath pacing, or cognitive engagement, embedding the neural shift into functional circuits.
Clinical reasoning: Treating the body means treating the brain. By choosing frequencies and targets that modulate specific central systems, we can transiently nudge networks toward healthier dynamics, then consolidate the state through behavior and repetition.
Entrainment and Resonance: Synchronizing Neural Networks for Functional Gain
Entrainment is the synchronization of oscillators through coupling. In neuroscience, coupling occurs via timed sensory inputs, motor rhythms, and autonomic cycles. When I deliver peripheral stimulation with precise timing, I can entrain distributed circuits—much like metronomes synchronizing on a shared platform.
- Conceptual model:
- Metronome analogy: Disparate oscillators (cortical columns, thalamic nuclei, autonomic cycles) synchronize when exposed to a consistent external driver. My stimulation acts as the driver; your neural nodes are the oscillators. Over time, coherence improves across networks.
- Physiologic resonance: Breathing frequency, muscle contraction rhythms, blood pressure waves, heart rate variability, and EEG bands (delta, theta, alpha, beta) exhibit cross-frequency coupling. Aligning interventions with intrinsic rhythms augments parasympathetic tone, cognitive clarity, and motor fluidity.
- “Frequency determines function”:
- 1–10 Hz: Emphasizes analgesia, endogenous opioids, and downregulation of hyperexcitability.
- 20–40 Hz: Enhances sensorimotor integration, vigilance, and attentional binding; useful for motor priming and rehabilitation.
- Timing around respiratory cycles can maximize vagal coupling; timing around movement onset can strengthen motor cortex entrainment.
Clinical reasoning: Entrainment provides a temporal handle on neural networks. Instead of only treating anatomical sites of dysfunction, I treat the time at which dysfunction arises—resetting reflexes and streamlining attentional resources. This approach can free cognitive bandwidth, improve postural control, and stabilize autonomic output.
Graph Theory, Neural Networks, and Clinical Decision-Making
The brain is not a simple serial chain; it is a graph—a system of hubs and edges operating in dynamic loops. This perspective explains why stimulating a single node can reconfigure distributed activity.
- Hubs and spokes:
- Think of Delta Airlines: hubs route traffic efficiently; the brain does the same with nuclei like the thalamus, basal ganglia, cerebellar deep nuclei, and prefrontal hubs.
- Edges connect hubs; strength and timing of edges determine functional connectivity.
- Neural networks and AI analogies:
- Modern AI models borrow from neural network concepts—layers, nodes, recurrent loops—because they reflect our brain’s parallel, distributed processing.
- Clinical takeaway: Adjusting one edge (e.g., median nerve input) can reweight connectivity across the graph, altering attention, motor readiness, and autonomic tone swiftly.
Clinical reasoning: I assess which hubs are underperforming (frontal timing, cerebellar accuracy, PMRF autonomic balance) and which edges need reinforcement (specific afferent lines). Then I select stimulation and manual interventions that strengthen those edges and re-balance hubs.
Practical Bedside Assessments: Frontal Lobe, Cerebellum, and PMRF
To translate theory into treatment, I rely on a triad of bedside tests that sample interconnected hubs:
- Frontal lobe rhythmicity and timing:
- Finger tapping: Informed by research on the Unified Parkinson’s Disease Rating Scale (UPDRS), rhythmic finger tapping assesses basal ganglia-thalamocortical timing. Slowness suggests basal ganglia/thalamus inefficiency; arrhythmia suggests cerebellar timing deficits; both reflect frontal integration.
- Saccadic latency: Instruct the patient to fixate your nose as you alternately wiggle a finger right or left. Observe the delay between the cue and eye movement. Side-specific latency differences indicate frontal eye field output and cerebellar vermis calibration variance.
- Cerebellar accuracy and postural control:
- Romberg: With eyes closed, sway direction often corresponds to ipsilateral cerebellar deficits. This test examines integrative balance with reduced visual compensation.
- Finger-to-nose, finger targeting: Accuracy—especially eyes closed—exposes cerebellar dysmetria. Fewer accurate touches and increased overshoot/undershoot on one side suggest hemispheric cerebellar mismatch.
- PMRF autonomic and extensor tone:
- Capillary refill asymmetry: Compress and release fingertip pads bilaterally; slower refill indicates greater sympathetic dominance on that side.
- Pupil responses: Light reflex speed and magnitude reflect autonomic balance and brainstem integrity.
- Muscle tone palpation: Evaluate extensor tone across limbs; PMRF contributes to extensor postural support and autonomic modulation.
- Blood pressure side-to-side: Differences can reveal asymmetric autonomic drive.
Clinical reasoning: I never treat based on a single finding. I connect the dots—frontal timing, cerebellar accuracy, and PMRF autonomic bias—then decide on side and modality. This detective mindset ensures safer, targeted interventions.
Functional Connectivity Mapping and Side Selection for Intervention
When the right hand shows slower finger tapping, I infer a left frontal lobe pacing issue. If Romberg sway is to the right, I suspect underperformance of the right cerebellum. If finger extensors are weaker on the left and autonomic signs suggest a left PMRF bias, I have a constellation: left frontal, right cerebellum, and left PMRF. This pattern shapes my intervention plan.
- Manual adjustment logic:
- Adjusting the right side of the body drives EPSPs to the left cortex, potentially improving left frontal timing and its downstream cerebellar links.
- The intention is to stimulate contralateral cortical activation via proprioceptive and nociceptive input.
- Electrical nerve stimulation side selection:
- Median nerve on the right sends potent efferents to the left thalamus and sensorimotor cortex, priming frontal circuits and binding sensorimotor representations.
- Pairing this with task practice (e.g., rhythmic tapping post-stimulation) consolidates gains.
- PMRF considerations:
- Since the frontal cortex projects ipsilaterally to PMRF for involuntary functions, improving left frontal output should normalize left PMRF tone—reflected in improved capillary refill, balanced extensor tone, and more stable autonomic signs.
Clinical reasoning: Select the side that best engages the underperforming contralateral cortex and the ipsilateral PMRF. Reinforce cerebellar accuracy via motor tasks and, when appropriate, cerebellar-targeted sensory stimulation (visual or vestibular tasks) to complement PNS.
Auricular Vagus and Trigeminal Stimulation: Pathways and Outcomes
- Auricular vagus nerve stimulation (aVNS):
- Pathway: Auricular branches → trigeminal nucleus → nucleus tractus solitarius → locus coeruleus → insula/anterior cingulate cortex.
- Outcomes: Increased parasympathetic tone, improved inflammation regulation, enhanced stress resilience, and potentially improved pain modulation via autonomic rebalancing.
- Practical tips: Use low to moderate intensities with careful electrode placement in auricular zones rich in vagal innervation; monitor heart rate variability and subjective calmness.
- Trigeminal stimulation:
- Pathway: Trigeminal distribution → trigeminal nucleus → thalamus → sensory cortex; collaterals to reticular formation.
- Outcomes: Boosted attention, arousal, and cortical excitability. Useful for post-concussive fatigue, attentional drift, and motor initiation challenges.
Clinical reasoning: Choose aVNS when autonomic imbalance, inflammation, or anxiety predominates; choose trigeminal stimulation to elevate alertness and cortical readiness. Both can be combined with breath entrainment or task initiation for a synergistic effect.
Median and Tibial Nerve Stimulation: Motor Priming, Pain Modulation, and Bladder Control
- Median nerve stimulation:
- Central effects: Activates dorsal column, thalamus, sensorimotor cortex; strong motor priming and cortical activation.
- Frequency selection:
- 20–40 Hz: Enhances sensorimotor binding and alertness—ideal for motor tasks, gait training, and preparatory activation in neurologic conditions.
- 1–10 Hz: Encourages endogenous opioid activity and analgesia; consider for pain-focused sessions or as a preliminary downregulation before motor practice.
- Clinical contexts: Tourette’s tic modulation via thalamocortical entrainment; concussion-related hypoarousal; Parkinson’s bradykinesia motor priming; Alzheimer’s attentional engagement.
- Tibial nerve stimulation:
- Central effects: Spinal afferents influence pain inhibition, endogenous opioids, and bladder control circuits.
- Evidence base: Significant research for incontinence and overactive bladder demonstrates central modulation via peripheral inputs. This is an elegant example of distal stimulation altering central autonomic outputs.
- Practical approach: Posterior tibial stimulation at conservative intensities, repeated sessions, and monitoring of voiding diaries, urgency scales, and HRV for autonomic changes.
Clinical reasoning: Use median nerve stimulation for motor system priming and attentional binding; use tibial nerve stimulation when targeting bladder control or analgesia through spinal-afferent-mediated central changes. Sequence matters: analgesic low-frequency sessions may precede motor priming at mid frequencies to reduce interference from pain.
Frequency Determines Function: Rationale for Parameter Selection
I select stimulation frequency based on desired neurochemical and network outcomes.
- Low frequency (1–10 Hz):
- Mechanisms: Favors mu-opioid receptors, descending pain inhibition, and LTD-like synaptic effects. Reduces hyperexcitable pain circuits, enabling relaxation and reconsolidation of normal sensory maps.
- Use case: Chronic pain dampening, pre-rehabilitation desensitization, sleep support adjuncts, and autonomic calming.
- Mid frequency (20–40 Hz):
- Mechanisms: Heightens sensory-motor binding, increases beta-range cortical readiness, improves attentional focus, and promotes short-term potentiation in motor circuits.
- Use case: Motor priming before gait training, upper-limb coordination tasks, cognitive tasks requiring sustained attention, and post-concussive activation.
- Timing and entrainment:
- Synchronizing with respiratory cycles can amplify vagal engagement.
- Aligning with movement initiation consolidates motor cortex plasticity.
Clinical reasoning: I often pair low-frequency analgesia early in a session with mid-frequency motor priming later, ensuring the patient is comfortable enough to engage but sufficiently activated to learn and retain movement patterns.
Building Durable Change: Pairing Stimulation with Behavior and Feedback
Immediate brain changes may not endure without structured reinforcement. To build durability:
- Task coupling: After stimulation, patients perform targeted motor tasks (e.g., rhythmic tapping, reach-to-touch accuracy drills) or attention drills (e.g., saccade training).
- Breath pacing: Slow diaphragmatic breathing (e.g., 6–8 breaths/min) promotes parasympathetic coupling, enhancing consolidation of calm network states.
- Feedback loops: Use HRV, reaction time, postural sway metrics, and subjective scales to inform session adjustments. Patients improve faster when we tailor inputs based on measurable responses.
Clinical reasoning: Neural entrainment creates a window of increased plasticity. Behavior performed during this window “writes” healthier patterns into the network, extending benefits beyond the session.
Autonomic Testing and Side-to-Side Asymmetries: Practical Measures
I routinely measure side-to-side autonomic differences:
- Capillary refill: A Delayed refill indicates sympathetic dominance and guides me to PMRF-related underperformance.
- Pupil light reflex: Speed and amplitude reflect autonomic balance; sluggish constriction can imply reduced parasympathetic vigor.
- Blood pressure differentials: May reveal systemic autonomic asymmetries, informing safer stimulation parameters.
- Skin temperature and sudomotor signs: Provide indirect readouts of sympathetic drive.
Clinical reasoning: These measures are quick, non-invasive, and anchor stimulation decisions to objective physiology, reducing the risk of overtreatment and guiding which side to stimulate or adjust.
Sensory-Motor Timing and Cerebellar Calibration: Detailed Rationale
The cerebellum calibrates timing, error correction, and precision. When finger tapping lacks rhythm, I suspect cerebellar contributions to frontal pacing. When eyes-closed targeting shows dysmetria, the cerebellar hemisphere likely needs reinforcement.
- Interventions:
- Pair median nerve stimulation with timed motor tasks to strengthen cerebellar-cortical coupling.
- Use visual fixation and vestibular-friendly stances to challenge balance while keeping tasks achievable.
- Consider dual-task paradigms (motor plus cognitive) for enhanced fronto-cerebellar integration.
Clinical reasoning: Cerebellar calibration benefits from consistent timing cues and repetitive, accurate practice just after stimulation, when plasticity peaks.
Protocol Design: Sequencing, Dosing, and Safety Checks
Designing a session requires thoughtful sequencing:
- Assessment:
- Baseline vitals, capillary refill, pupil responses, muscle tone, finger tapping rhythm, saccadic latency, Romberg sway, targeting accuracy.
- Priming:
- Manual adjustment on the side selected to activate the contralateral cortex, with conservative dosing and patient feedback.
- Electrical Stimulation:
- Choose nerve and frequency:
- Pain or autonomic downregulation: 1–10 Hz (tibial or auricular vagus).
- Motor priming and alertness: 20–40 Hz (median nerve, trigeminal zones).
- Dosing: Start with short bouts (e.g., 2–5 minutes), low to moderate intensity; reassess.
- Choose nerve and frequency:
- Task Pairing:
- Targeted motor or cognitive drills aligned with identified deficits.
- Reassessment:
- Repeat key measures to confirm direction of change and adjust parameters accordingly.
- Documentation:
- Record side selection rationale, frequencies, intensities, patient sensations, and objective changes.
Safety checks: Monitor for adverse sensations, autonomic instability, or unexpected fatigue. In patients with communication barriers, keep dosing minimal and use physiologic markers to guide safe titration.
Avoiding Single-Finding Therapy: The Medico-Legal and Ethical Imperative
Treating based on a single sign invites error. I teach clinicians to think like detectives: gather multiple convergent findings before acting. In the United States—an intensely litigious environment—each treatment is a professional risk decision. Mitigate risk by building strong cases: multiple tests, documented rationale, objective outcomes, and patient-informed consent.
Clinical reasoning: When the “hand in the deck” is visible—robust data pointing in the same direction—your therapeutic bet is safer. If findings conflict, pause and reassess rather than forcing a protocol.
Communication-Limited Patients: Ethical Caution and Practical Steps
For patients unable to report discomfort (e.g., severe brain injury), I model the stimulation on myself first using identical parameters. I then apply minimal intensities, continuously observe autonomic signs, and use short bouts with frequent checks. When in doubt, defer intervention until more robust monitoring or specialist coordination is available.
Clinical reasoning: Absence of communication makes adverse events harder to detect. Ethics demand extra caution, conservative dosing, and interdisciplinary oversight.
Integrating Hydrogen Supplement Anecdotes with Evidence-Based Care
Occasionally, patients report anecdotal benefits from supplements such as hydrogen water. While some literature suggests antioxidant effects, I avoid generalized recommendations—especially in pregnancy—without coordinated medical guidance. Anecdotes do not replace trials; I focus on modalities with stronger evidence and always defer to obstetric or primary care recommendations for pregnant individuals.
Clinical reasoning: Distinguish curiosity from clinical certainty. Keep anecdotes in perspective and anchor care in reproducible, safety-oriented methods.
Documentation, Consent, and Long-Term Strategy
- Documentation:
- Capture pre/post assessments, stimulation parameters, side selection rationale, and any adverse events.
- Note patient education, consent, and session goals.
- Long-term strategy:
- Begin with conservative dosing; progress based on demonstrated tolerance and objective improvements.
- Cycle frequencies: analgesia before activation when pain interferes; activation before task learning when attention is the limiting factor.
- Encourage home routines: breath pacing, rhythmic movement drills, sleep hygiene, and hydration to support autonomic balance.
Clinical reasoning: Clear records improve continuity, safety, and medico-legal protection. Patients who understand the “why” of care are more adherent and gain better outcomes.
Putting It All Together: A Step-by-Step Clinical Template
- Identify hubs and edges:
- Frontal timing (tapping, saccades)
- Cerebellar accuracy (Romberg, finger-to-nose)
- PMRF autonomic bias (capillary refill, pupils, BP)
- Select intervention side and modality:
- Manual priming on the side that activates the underperforming contralateral cortex.
- Electrical stimulation targeted to the chosen labeled line (median, tibial, trigeminal, auricular, and vagus).
- Choose frequencies:
- 1–10 Hz for analgesia and autonomic downregulation.
- 20–40 Hz for motor priming and attentional binding.
- Entrain and consolidate:
- Pair with breathing or movement at synchronized tempos.
- Immediate task practice to harness primed plasticity.
- Reassess and iterate:
- Recheck the triad measures; adjust dosing or sides as needed.
- Safety gate:
- Screen contraindications; monitor; defer when uncertain.
The Brain’s Core Purpose: A Continuous Feedback Loop
As a clinician deeply invested in the neurophysiology behind healing, I often find myself returning to a foundational question: Why do we have a brain? The answer, in its elegant simplicity, is the key to understanding the profound impact of therapies like chiropractic. Our brain exists to perform a continuous, vital cycle: it receives information from our environment, interprets that information, and then formulates and executes an appropriate response. This entire cycle is what allows us to survive, adapt, and thrive. When this cycle is functioning optimally, we experience health. When it breaks down, we experience dysfunction and disease.
Let’s visualize this process. In a healthy individual, our vast network of sensory receptors—in our skin, muscles, joints, eyes, and ears—is constantly gathering data. They are our biological antennas. This information travels as electrical signals along neural pathways to the brain. The brain, our central processor, interprets this flood of data in the context of our memories, goals, and current state. It then sends out commands to our muscles and glands, directing them to act. “You felt this, so do this.” The muscles execute the command, and in doing so, they create new sensory information—feedback—which is then sent back to the brain by the receptors. It’s a beautiful, self-regulating loop. When everything is working correctly, information flows in, is processed, a response is generated, and the outcome is fed back into the system seamlessly. This is the signature of a healthy, adaptive nervous system.
When the Body Is Injured: Garbage In, Garbage Out
Now, let’s consider what happens when there’s an injury to the body. This could be anything from a classic spinal subluxation, where a joint is restricted in its movement, to a frozen shoulder, a sprained ankle, or even the immobilization of a limb in a cast. The fundamental problem is the same: the body is not working properly.
In this scenario, the brain still sends out its commands. It might tell the shoulder, “Lift the arm.” However, due to the injury—adhesions, inflammation, and joint fixation—the arm cannot move as instructed. The mechanoreceptors within the muscles and joints of that shoulder, which are designed to report on movement, position, and stretch, are now sending back distorted, corrupted information. They are essentially telling the brain, “We’re not moving,” or “The movement is painful and restricted,” even when the initial command was for normal motion.
This is the “garbage in” part of the equation. The brain receives this aberrant sensory data. Because the brain’s output is based on its input, it starts generating faulty motor commands in response—the “garbage out.” It might try to compensate by recruiting other muscles that aren’t designed for the job, leading to strain and further injury. It might create a state of chronic muscle splinting and guarding around the injured area. This faulty motor output creates even more bad sensory feedback, which is then re-consumed by the receptors. A vicious cycle is established. The initial physical problem has now created a neurological dysfunction, a cycle of bad information that perpetuates pain, restriction, and poor function. This principle—garbage in, garbage out—is a fundamental law of information processing, as true in software engineering as it is in human neurobiology.
When the Brain Is Injured: A Breakdown in Processing
The feedback loop can also be disrupted from the other direction: when the brain itself is injured or dysfunctional. A poignant example is found in individuals with certain neurodevelopmental conditions like autism spectrum disorder, or in cases of traumatic brain injury such as a concussion.
In many of these cases, there is nothing inherently wrong with the peripheral sensory receptors. The photoreceptors in the retina are detecting light perfectly. The mechanoreceptors in the skin are accurately sensing touch. The auditory receptors in the ears are faithfully converting sound waves into neural signals. The information coming in is perfectly valid and accurately represents the external environment.
However, the problem lies in the central processor—the brain. Due to differences in neural wiring, inflammation, or metabolic dysfunction, the brain cannot interpret this sensory information correctly. A normal level of auditory input might be perceived as overwhelmingly loud and threatening. A light touch might feel noxious. The brain, misinterpreting the data, then generates an output that appears inappropriate for the situation. This can manifest as hand-flapping, covering the ears, or a heightened startle response.
These motor actions, while a logical response from the brain’s distorted perspective, are not adaptive for survival in the real world. Furthermore, these actions generate their own set of sensory feedback. The receptors now report this unusual motor activity back to the already struggling brain. This new stream of “bad” information further reinforces the dysfunctional processing, creating a downward spiral of neurological dysregulation. This is the core of my theoretical model for understanding the long-term consequences of concussion—an initial injury to the processor creates a self-perpetuating cycle of sensory-motor dysfunction.
This fundamental concept, the Safety Pin Cycle, was articulated by the founders of chiropractic, the Palmers, back in the 1880s. They depicted the brain sending a signal down to a tissue cell and the tissue cell sending a signal back up to the brain, forming a closed loop. A subluxation was seen as the “unclasped” pin, interrupting that vital flow of information. They were truly ahead of their time, describing a core principle of systems neuroscience long before the tools existed to fully validate it. Their insight remains profoundly relevant today as we explore the neurological basis of health and healing.
The Fundamental Purpose of the Brain: The Sensorimotor Loop
If you thought I was excited about PBM, this is truly my favorite part of the discussion. The answer to why we have a brain is surprisingly simple and profoundly elegant.
Our brain devotes the vast majority of its volume, energy, and computational power to processing various sensory inputs from the body in order to determine and initiate appropriate, coordinated motor outputs to the body.
In short: The only reason we have a brain is to move or not to move.
This applies to everything. The movement of our arms and legs is obvious. But it also applies to the beating of our heart, the secretion of saliva from our parotid glands, or the release of cortisol from our adrenal glands. These are all “motor” actions controlled by the brain—an instruction to do something or not to do something.
The scientist who brilliantly spearheaded this philosophy is Dr. Daniel Wolpert. He uses the perfect biological example to illustrate this point: the sea squirt.
A sea squirt begins its life as a tadpole-like creature with a very primitive nervous system. It uses this nervous system to propel itself through the water, sucking water in and spitting it out to move. It swims until it finds a suitable rock or piece of coral to call home. Once it finds its permanent spot and attaches itself, the very first thing it does is consume its own nervous system for food. It digests its own brain and spinal cord. Why? Because it will never move again. It becomes a stationary filter feeder for the rest of its life, and a nervous system is a metabolically expensive organ to maintain if you don’t need it. The sea squirt is the ultimate proof that the nervous system exists solely for the purpose of movement.
The Sensorimotor Loop: The Blueprint of a Functioning Nervous System
I have simplified this profound concept into a diagram that I call the Sensorimotor Loop. This loop is the basis of all neurological function.
(Note: Representative image description.)
There is no official starting point, but let’s begin with sensory input.
- Sensory: This is the transduction of an environmental stimulus into an action potential. Transduction simply means converting one form of energy into another. The pressure on your skin, the light hitting your retina, the sound waves vibrating your eardrum—all of these are converted into the electrical language of the nervous system.
- Integration: The sensory information travels up to the brain, where it is integrated. Integration is the confluence and comparison of current and past environmental potentials. Your brain takes the incoming sensory data and asks, “What is this? What does it mean? Have I experienced this before? What happened last time?” It compares the current state to a vast library of past experiences to make sense of the world.
- Motor: Based on that integration, the brain generates a motor output. The definition of “motor” is the imparting or production of motion or action. This is the brain’s decision, executed as a command to the body.
Crucially, inaction is also a form of action. If you have an impulse to say something hurtful but you stop yourself, that inhibition—that “no-go” signal—is an active motor command from your prefrontal cortex. It required brain energy to stop the action.
- Re-entry as Sensory Input: The loop doesn’t end there. As soon as you perform a motor action (or inaction), you immediately sense the consequences of that action. If I feel an itch on my neck (sensory), I integrate that information and decide to scratch it (motor). As my fingers move across my skin, I am sending new sensory information back to my brain about the pressure and movement of my hand and whether the itch is going away. My brain integrates this new information and says, “The itch is gone.” The next motor output is then an inaction: “Stop scratching.”
This continuous, recursive loop is how our brain works, moment to moment, for our entire lives.
This very concept forms the basis of a scientific paper I published in January of 2026. It presents, for the first time, a comprehensive theory on why persistent post-concussion symptoms occur. I’m quite proud of the catchy name I came up with for the model: NERD, which stands for Network Entrapment by Reflex Dysfunction. If you’re interested in a deep, heavy read, I encourage you to look it up—you can simply go to Google and search for “antigenic nerd,” and it should be the first result.
When you read that paper, you will see how this Sensorimotor Loop is at the very heart of the theory. And you will also see how chiropractic care, which fundamentally intervenes in this loop by providing powerful mechanoreceptive (sensory) input and restoring motion (motor function), fits perfectly within this model of neurological healing.
By understanding this loop, we understand the blueprint for rehabilitation. A problem can arise from faulty sensory input, faulty integration, or faulty motor output. Our job is to identify where the loop is broken and use targeted therapies—whether it’s photobiomodulation, chiropractic adjustments, vestibular rehabilitation, or cognitive exercises—to repair it.
Sensory Transduction: Turning the Physical into the Neurological
To truly grasp how a chiropractic adjustment or any sensory-based therapy works, we must first understand the magical process of sensory transduction. In simple terms, transduction is the conversion of energy from one form into another. This happens all around us. A battery converts chemical energy into electrical energy. A solar panel converts photonic (light) energy into electrical energy. A microphone converts the mechanical energy of sound waves into electrical energy.
Our bodies are masters of transduction. Your ability to hear my words right now is a perfect example. Sound waves, which are mechanical vibrations in the air, strike your eardrum, causing it to vibrate. A series of tiny bones in your middle ear amplifies these vibrations and transmits them to the fluid-filled cochlea in your inner ear. Inside the cochlea, tiny hair cells bend in response to the fluid waves, and this physical bending opens ion channels. The resulting flow of ions creates an electrical signal—the language of the brain. You have just transduced mechanical energy into electricity.
When I perform a chiropractic adjustment, I am applying a controlled, high-velocity, low-amplitude mechanical force to a joint. This force rapidly stretches the joint capsule and the surrounding muscles. This physical stretch activates a class of sensory receptors called mechanoreceptors, which are specifically designed to detect mechanical pressure, vibration, and stretch. They are the primary transducers for manual therapy. The mechanical energy of the adjustment is instantly converted into a burst of electrical signals that surge toward the spinal cord and brain.
Similarly, other therapies utilize different forms of transduction. If we were to use a modality like iontophoresis, which uses an electrical current to drive medication through the skin, or if we administer a medication orally, we are engaging chemoreceptors. These receptors respond to specific chemical molecules, binding to them and initiating an electrical signal. In this way, chemical energy is transduced into the electrical currency of the nervous system.
The Receptors: Our Body’s Sensor Network
Our sensory transducers can be broadly categorized based on the information they gather:
- Exteroceptors: These receptors provide information about the external environment. They include the photoreceptors in our eyes, the auditory receptors in our ears, and the various receptors in our skin for touch, pressure, temperature, and pain.
- Interoceptors: These receptors monitor the internal state of our body. They are found in our organs and blood vessels and report on things like blood pressure, oxygen levels, and visceral distension.
- Proprioceptors: This is a crucial category for understanding manual medicine. Proprioceptors provide information about the position and movement of our own bodies. They are the mechanoreceptors located in our muscles (muscle spindles), tendons (Golgi tendon organs), and joint capsules. They are what allow you to touch your nose with your eyes closed.
For our discussion today, we are primarily concerned with mechanoreceptors, as they are the main targets of a chiropractic adjustment. However, it’s vital to recognize that direct nerve stimulation, which we will also discuss, bypasses the receptor and activates the nerve fiber itself, offering another powerful therapeutic pathway.
The Language of Receptors: Graded Potentials
Now, let’s dive into a critical concept from neurophysiology that is absolutely essential for understanding how therapies can be dosed and tailored to individual needs: the difference between a receptor potential and an action potential. This may bring back some memories (or PTSD) of neurophysiology class, but I promise it’s clinically relevant.
An action potential is the signal that travels long distances down a nerve axon. It is famous for its “all-or-none” property. This means that once the neuron’s threshold is reached, it fires with a fixed, maximum intensity. It either fires at 100% or it doesn’t fire at all (0%). It’s like a light switch—it’s either on or off. Furthermore, action potentials are propagated and non-degrading. Once initiated, they travel the full length of the axon without losing strength, ensuring the signal arrives at its destination intact.
Receptor potentials, however, are completely different, and this difference is everything. Receptor potentials are the initial electrical events that happen at the sensory receptor itself when it’s stimulated. They have two key properties that stand in stark contrast to action potentials:
- They are Graded: This is the most important property for us as clinicians. “Graded” means that the size of the receptor potential is directly proportional to the intensity of the stimulus. It does not follow the all-or-none rule. Instead of being 0% or 100%, it can be 1%, 10%, 50%, or any value in between. This means a soft, gentle adjustment produces a small receptor potential. A medium-force adjustment produces a medium-sized potential. A strong, high-force adjustment produces a large potential. These are three neurologically distinct events. The brain receives a different message depending on the force, speed, and amplitude of the adjustment. This is why technique matters. This is why sometimes a patient needs a gentle, low-force mobilization, and other times they require a robust, high-velocity thrust. We are modulating the “volume” of the neurological input based on the patient’s specific needs.
- They are Non-Propagated: This means the receptor potential is a local event, confined to the area of the receptor. It doesn’t travel down the nerve fiber. Think about it: touch your finger to your arm. You feel the sensation. Now, take your finger away. The sensation is gone instantly. The receptor stops firing the moment the stimulus is removed. This is because the receptor potential is non-propagating. It only exists as long as the stimulus is present. This is incredibly powerful. It means that the neurological stimulation from a chiropractic adjustment is discrete and controllable. The massive burst of efferent input occurs during the thrust, and then it’s over. We are not creating a runaway, continuous signal. We are providing a precise, powerful, and temporary burst of information to reset the system.
These graded, non-propagated receptor potentials trigger what are known as Excitatory Postsynaptic Potentials (EPSPs) at the first synapse in the spinal cord. And this is where the real magic begins.
From Adjustment to Gene Expression: The Cellular Cascade
If you want to understand how chiropractic works on a fundamental, biological level, this is the single most important concept we will discuss. When someone, even a well-meaning colleague like an orthopedic surgeon, dismisses chiropractic by saying, “Oh, it just releases endorphins,” they are missing the vast, intricate, and profound cellular story that unfolds with every single adjustment.
I recently had a candid conversation with a good friend of mine, a brilliant orthopedic surgeon. We have a great relationship, so we can talk honestly about our professions. I asked him, “What do you honestly think chiropractic does? Not what you were taught in medical school—which he admitted was virtually nothing, aside from warnings about vertebral artery dissection—but what do you, as a scientist, think it does?” He paused and said, “I can’t really fathom why cracking a bone would do anything for you… Maybe your body releases endorphins?”
His response, using the phrase “cracking a bone,” is telling. It highlights a massive communication and education gap between our professions. But his hypothesis about endorphins is a reasonable starting point. Endorphins are powerful neuropeptides that can reduce pain, elevate mood, and increase energy. An adjustment certainly can trigger their release. But that is merely the tip of the iceberg. The reality is infinitely more complex and impactful. Even within the chiropractic profession, there is confusion. In an online discussion, I once posed the question, “What are we actually adjusting?” One chiropractor replied emphatically, “We adjust bones!” I countered that I believe we adjust joints. His retort was, “Have you ever touched a joint in your life?” He was being semantic, of course—I am touching skin that overlies muscle that overlies a joint capsule that contains the bone. But the exchange reveals that we often lack a unified, precise language to describe the neurological event we are initiating.
So, let’s build that language. Let’s map out what is actually happening from a neurological basis.
The Power of the EPSP
As we established, the mechanical force of an adjustment is transduced by mechanoreceptors into graded receptor potentials. These potentials cause the sensory neuron to release neurotransmitters at its synapse with the next neuron in the chain, typically in the dorsal horn of the spinal cord. This creates an Excitatory Postsynaptic Potential (EPSP). The name tells you exactly what it does: it is a potential that has the ability to excite the postsynaptic (receiving) neuron.
These EPSPs are chemical messages that diffuse across the synapse and bind to receptors on the receiving neuron. This binding opens ion channels, causing a small, localized depolarization of the neuron’s membrane. A single EPSP is usually not enough to make the neuron fire an action potential, but the massive, synchronized burst of input from a chiropractic adjustment generates a huge summation of EPSPs, powerfully activating these pathways.
But the EPSP does more than just contribute to firing an action potential. It initiates a cascade of events inside the cell. Here is the step-by-step process:
- Enzyme Activation: The influx of ions (such as calcium) and the binding of neurotransmitters activate a host of intracellular enzymes, particularly kinases. Kinases are “master switches” in the cell; their job is to phosphorylate other proteins, turning them on or off.
- Immediate-Early Gene Response: These activated kinases trigger a Cellular Immediate-Early Gene (IEG) Response. This is a rapid, transient activation of a specific set of genes that act as first responders to cellular stimulation. Think of them as the supervisors who are called in to manage a new project. The best-studied of these IEGs are proto-oncogenes, including c-Fos, c-Jun, and c-Myc. Now, don’t let the term “proto-oncogene” scare you. While their uncontrolled expression is linked to cancer, in a healthy neuron, they are essential regulators of cell growth, differentiation, and adaptation. Their presence is a measurable, scientific marker that a neuron has been stimulated. This is not chiropractic theory; this is fundamental molecular biology. The dots we are connecting are simple: chiropractic stimulates receptors, receptors create EPSPs, and EPSPs are known to induce c-Fos expression.
- Mitochondrial Activation: The IEG response sends a signal to the cell’s power plants: the mitochondria. They get the message: “Gear up! We’re about to do some heavy lifting.” The mitochondria ramp up production of ATP and GTP, the energy currencies of the cell. This energy is essential for the next, most resource-intensive steps.
- Transcription and Translation: Now, with the energy supply secured, the real work begins. The IEG proteins act as transcription factors. They travel into the cell’s nucleus, bind to the DNA, and initiate the process of transcription. This is where a specific segment of DNA is “read” and a complementary copy is made in the form of messenger RNA (mRNA). Why? Because the DNA, the master blueprint, is too precious to leave the protected environment of the nucleus. The pores in the nuclear envelope are too small for it to exit. So, a disposable mRNA copy is made.
- Protein Synthesis: The mRNA molecule leaves the nucleus and travels to the endoplasmic reticulum (ER) in the cell’s cytoplasm. Here, in structures called ribosomes, the process of translation occurs. The genetic code on the mRNA is read, and a specific protein is synthesized, amino acid by amino acid. This is the ultimate goal of the entire cascade: protein synthesis.
The Proteins of Neuroplasticity
What kinds of proteins are we making? We’re making everything the neuron needs to maintain itself, repair itself, and adapt. This includes:
- Structural Proteins: Things like tau proteins and actin form the neuron’s internal cytoskeleton, the scaffolding that gives the cell its shape and stability. They are like the studs and beams of the cellular hotel, keeping it structurally sound.
- Neurotransmitters: The very chemical messengers that allow neurons to communicate with each other are proteins or are synthesized by protein-based enzymes.
- Receptors: The proteins that sit on the cell surface and receive incoming signals are constantly being replaced and regulated.
- Proteins for Neuroplasticity: This is where it gets truly exciting. The cell synthesizes proteins like Brain-Derived Neurotrophic Factor (BDNF), a master molecule for neuronal growth, survival, and the formation of new connections (synapses). It’s literally fertilizer for the brain.
This entire process, from the EPSP to the creation of new proteins, is the biological basis of neuroplasticity. It is how our nervous system changes and adapts in response to experience. Every single time you deliver an adjustment, you are not just moving a bone. You are initiating a gene-expressing, protein-synthesizing, neuroplastic event.
And here’s a crucial point: this is not unique to chiropractic. Shining a light in someone’s eye does the same thing in the visual cortex. Playing sound in their ears does the same thing in the auditory cortex. Direct nerve stimulation does the same thing along the targeted pathway. The mechanism—EPSP leading to protein synthesis—is universal. The difference lies in the receptors we stimulate, the pathways we activate, and the areas of the brain we target. Chiropractic’s unique power lies in its ability to deliver a massive, synchronized input to the proprioceptive system, which has vast and powerful connections throughout the entire brain and central nervous system.
It’s also important to note that many of the proteins we hear about in the context of neurodegenerative disease—like tau, alpha-synuclein (associated with Parkinson’s), and APOE (a cholesterol transport protein linked to Alzheimer’s)—are normal, necessary proteins. Disease occurs not because these proteins exist, but when their production becomes dysregulated or, more often, when the cellular clearance mechanisms that are supposed to remove old, misfolded proteins fail. By stimulating the neuron’s healthy maintenance and repair processes, we may be helping to keep this delicate balance in check.
Finally, all of these newly synthesized proteins and neurotransmitters are packaged up by the cell’s “shipping department,” the Golgi apparatus, and transported down the axon to be released at the next synapse, ready to influence the next neuron in the chain. An EPSP is, therefore, a neuron-generating and neuron-sustaining event. It is the fundamental unit of neural health and adaptation.
From Cellular Activity to Brain-Wide Arousal: The ARAS
We’ve seen how a single adjustment can trigger a profound cascade of activity within individual neurons. Now, let’s zoom out and look at the bigger picture. What happens when millions of these neurons are firing together, sending their signals upstream into the brain?
This question brings us to the pioneering work of Sir Charles Sherrington, who, in 1906, received the Nobel Prize for his discoveries. He described a critical network in the brainstem that he named the Ascending Reticular Activating System (ARAS). The ARAS is a diffuse network of neurons that acts as the brain’s master arousal center and sensory filter.
Sherrington defined the ARAS as the nexus where the sum of all excitatory and inhibitory impulses from the body converges. Every piece of sensory information from your skin, muscles, joints, eyes, and ears sends collateral branches into this system on its way to the cerebral cortex. ARAS’s job is to take this cacophony of incoming data and decide what is important enough to bring to conscious awareness. It is the gatekeeper of consciousness.
All of this sensory input funnels into a critical structure deep in the brain: the thalamus. The thalamus is often called the “sensory relay center,” which is true, but that description dramatically undersells its importance. The thalamus is a complex integrator for virtually all cortical activity—sensory, motor, cognitive, and emotional. It’s the grand central station of the brain.
The constant, massive flow of sensory information from the body into the ARAS and thalamus generates the thalamic resonant rate, or thalamocortical rhythm. This is essentially the background electrical “hum” of the brain, a rhythmic firing that keeps the cerebral cortex awake, alert, and ready to process information. Think of it as the brain’s idle speed. A healthy, robust sensory input from the body, particularly from the large-diameter proprioceptive fibers in the spine and limbs, creates a strong, stable thalamocortical rhythm, leading to an alert, focused, and healthy brain. When sensory input is diminished or corrupted—as in cases of chronic pain, joint fixation, or sensory deprivation—this rhythm can become destabilized, leading to issues like brain fog, fatigue, and poor cognitive function.
This brings us full circle, connecting modern neuroscience back to the earliest days of chiropractic. Let’s rewind to the late 1800s and the famous story of Harvey Lillard. He reportedly lost his hearing after feeling a “pop” in his back while bending over. D.D. Palmer examined him, found a displaced vertebra, delivered an adjustment, and Lillard’s hearing was restored. Whether you take the story literally or as an allegory, its core insight is profound. It suggests that a mechanical issue in the spine can have far-reaching functional consequences and that a mechanical correction can restore that function.
From the perspective of the ARAS, it’s a plausible neurological mechanism. The massive proprioceptive and nociceptive input from the spine feeds directly into the brainstem and ARAS. A significant disruption (the subluxation) could theoretically alter the baseline firing of this system, potentially impacting the processing of other sensory modalities, like hearing. The adjustment, by delivering a powerful, normalizing burst of proprioceptive input, could have rebooted the ARAS, restoring its normal function and allowing for proper sensory processing once again. This is the power of understanding the nervous system not as a collection of separate parts, but as a deeply integrated, holistic system.
EPSPs from Manual Adjustments: Priming Thalamocortical Networks
Manual interventions—particularly adjustments and precise mobilizations—can elicit excitatory postsynaptic potentials (EPSPs) that propagate through thalamocortical circuits. This aligns with functional neurology findings: appropriately dosed proprioceptive inputs can transiently improve cortical excitability, motor readiness, and attention.
- Mechanism:
- Proprioceptive afferents engage dorsal columns and brainstem relays, arriving at the thalamus and sensorimotor cortex.
- Adjustments performed on the right side favor activation of the left cortex, and vice versa. This supports side selection guided by bedside testing (e.g., left frontal underperformance → right-sided adjustment to drive contralateral cortex).
- Integration:
- I often combine adjustments with carefully timed median nerve stimulation to reinforce the EPSP-driven priming effect.
- Immediately following, I introduce motor tasks that leverage the primed state, improving carryover.
Clinical reasoning: The goal is sequence synergy—manual priming, electrical entrainment, and task consolidation—to yield meaningful functional gains.
Neurophysiology, Perception, and the Central Integrative State: How Spinal Input Shapes Brain Function
In my clinical practice, I often encounter scenarios where patients report sudden changes in perception—such as diminished hearing, blurred visual clarity, or muted proprioception—following a mechanical event like a fall. A traditional storyline might claim that a “bone impinging on a nerve” is responsible, blocking the flow of innate intelligence. While this imagery had pedagogical appeal in the early evolution of manual healthcare, the modern neurophysiological explanation is more precise and far more powerful.
The concept that explains this clinically is the central integrative state—the dynamic readiness, responsiveness, and connectivity of cortical networks, driven in part by cerebellar-thalamocortical loops. The cortex does not operate in isolation; it is continuously modulated by ascending sensory input and descending predictions. Proprioceptive signals from spine and limb joints—especially fast-adapting mechanoreceptor input—feed the cerebellum and thalamus, which in turn regulate cortical firing thresholds. When a spinal region becomes hypomobile, mechanoreceptive traffic can decrease or distort, leading to reduced excitatory postsynaptic potentials (EPSPs) streaming into thalamic relay nuclei. As EPSP volume declines, thalamic gating stiffens, cortical networks drop below perceptual threshold, and the person may transiently “lose” aspects of perception—be it sound clarity, positional sense, or nuanced somatosensory detail. This is a systems-level shift—not mechanical impingement—explaining why symptoms can resolve when mobility and afferent patterning are restored.
Importantly, a clinical intervention that reintroduces robust, well-timed afferent input can ramp EPSPs, re-open thalamic gating, and restore the central integrative state. Manual techniques, sensorimotor drills, electrical neuromodulation, and vestibular-ocular stimuli can each, through different mechanistic routes, elevate network responsiveness and bring the system back above threshold. The reason these diverse tools can produce convergent outcomes is that their common denominator is afferent-driven modulation of thalamocortical dynamics.
The Physiological Blind Spot: A Window into Thalamocortical Function
Clinically, one of the most elegant low-tech measures of cortical-thalamic function is the physiological blind spot. Anatomically, the optic nerve head enters the retina at a site devoid of photoreceptors, producing a literal gap in retinotopic input. Yet most of us do not “see” a hole in our vision because the brain actively fills it via predictive coding and contextual interpolation. The size of the perceptual blind spot is not merely anatomical—it is modulated by the integrity and activity of thalamocortical networks.
When cortical networks are robust—high central integrative state—the brain’s capacity to interpolate content within the blind spot is greater, effectively shrinking the perceived area. Conversely, when cortical activity is depressed or dysregulated, the blind spot often enlarges. This phenomenon becomes a practical biomarker: if the blind spot measured monocularly is larger on the right visual field, this suggests a relative functional deficiency of the contralateral (left) thalamocortical apparatus. Because somatosensory afferents from the right side of the body preferentially project to the left thalamus, one can leverage side-specific interventions to modulate asymmetries.
In clinical application, I carefully map each patient’s blind spot—connecting peripheral disappearance points to calculate area—and compare right versus left. This measurement is repeatable and, while not diagnostic in isolation, it offers a tangible index of thalamocortical balance. Using this information, I design side-specific proprioceptive interventions, manual input, or neuromodulation parameters to target the underactive contralateral network.
From Debate to Data: Why “Pressure on a Cranial Nerve” Is Not the Best Model
I vividly remember confronting the dissonance between simplistic explanations—“a bone pressing on a nerve reduces flow of innate intelligence”—and formal neuroanatomy. For instance, claims that thoracic segments directly “compress cranial nerve VIII” (the vestibulocochlear nerve) do not align with anatomy: cranial nerve VIII arises from the brainstem and traverses the internal auditory canal; it is not anatomically subject to thoracic compression. Yet, paradoxically, patients sometimes experience altered hearing after spinal injury or immobility. How can this be reconciled?
The reconciliation lies in network physiology. Auditory perception depends on the coherent activity of brainstem centers, thalamic relay (medial geniculate nucleus), and auditory cortices, influenced by multi-sensory integration, attentional systems, and cerebellar modulation of timing. If spinal immobility reduces somatosensory drive, thalamic readiness for processing incoming auditory signals can decline. The system falls below perceptual threshold—not from local crush—but from reduced excitatory gain within thalamocortical loops. Restoring movement and mechanosensory input, whether via manual therapy or patterned stimulation, can elevate EPSPs, re-open thalamic gating, and improve auditory clarity. In other words, hearing shifts emerge from centrally mediated state changes, not peripheral compression of cranial nerve VIII by distant bones.
This modern framework clarifies inconsistencies that historically fueled philosophical debate. It honors clinical observations while grounding them in evidence-based neurophysiology.
Evidence of Brain Changes Following Cervical Manipulation: Modern Research Perspectives
Research has shown that cervical spinal manipulation can alter measures of brain function. Early work proposed that manipulative inputs affect thalamocortical dynamics, and modern studies have used objective tools—such as EEG, evoked potentials, fMRI, and functional near-infrared spectroscopy—to evaluate changes in cortical activity and sensorimotor integration following spinal interventions. Additionally, sophisticated assessments like blind spot mapping provide a simple but insightful correlate of central state modulation.
Key methodological points that strengthen modern interpretations include:
- The use of standardized protocols and randomization where appropriate.
- Objective pre- and post-intervention measures, including visual field tests, cortical oscillatory analysis, or sensorimotor performance metrics.
- Careful side-specific dosing to test hypotheses about contralateral thalamic modulation.
- Recognition that effect direction (e.g., blind spot shrinking versus expanding) must be interpreted relative to baseline function: a change is an effect; whether it is beneficial depends on context.
These approaches underscore that spinal manipulative therapy is not merely about joint motion but about afferent input engineering—crafting sensory experiences that reshape cortical responsiveness.
Cerebellar-Thalamocortical Loops and the “Volume” of Perception
At the heart of this discussion lies the cerebellar-thalamocortical loop, a continuous circuit where cerebellar outputs modulate thalamic relay fidelity and cortical synchronization. The cerebellum is not only a motor coordinator; it is a timing and prediction engine for both sensorimotor and cognitive domains. When proprioceptive input from the spine and limbs is rich and well-timed, the cerebellum can refine thalamic gating, enabling cortical networks to maintain high integrative states. Conversely, degraded input diminishes thalamic relays, slowing cortical oscillations and weakening network coherence.
Three physiological principles are crucial:
- EPSP Density and Timing: High-frequency, well-patterned afferent signals increase EPSP density, summating to push neural assemblies above response thresholds. Timing matters; chaotic input may increase energy without improving coherence.
- Oscillatory Synchronization: Thalamocortical systems rely on rhythmic loops. Frequencies such as gamma (high-frequency engagement) or alpha (idling/inhibitory) modulate attentional gating and sensory prioritization. Targeted sensory tasks or electrical stimulation can bias frequency bands.
- Plasticity and Network Re-tuning: Repeated, well-structured input fosters synaptic plasticity—changes in synaptic strength and network architecture that sustain improved function beyond the immediate intervention window.
Clinical interventions aim to “turn up the volume” of useful signals and promote the rhythmic synchronization that supports clear perception and efficient behavior.
Altered Oscillations, Altered Reality: Neurofeedback and Afferent Engineering
If the frequency of oscillation and network synchronization shape what we perceive, then any intervention that shifts oscillatory patterns can change a person’s subjective reality. Tools like neurofeedback explicitly train cortical rhythms, using operant conditioning to increase or decrease activity in targeted bands and regions. Similarly, manual therapy, vestibular challenges, visual-oculomotor drills, and electrical neuromodulation alter afferent input and downstream oscillatory behavior.
For example:
- A patient with anxiety may exhibit excessive high-beta activity in certain networks. Interventions that encourage alpha or theta coherence can reduce hypervigilance.
- A person with attentional dysregulation may benefit from gamma-enhancing tasks (e.g., fast-paced visuomotor challenges) combined with side-specific proprioceptive inputs to boost thalamocortical readiness.
The practical takeaway is profound: sensory engineering can reshape internal experience. Yet this power requires responsibility. We must tailor interventions to the individual’s baseline state and goals, monitor objective markers, and avoid indiscriminate stimulation that could disrupt useful network patterns.
“Noise In, Noise Out”: A Unifying Principle Across Disciplines
Across chiropractic, osteopathy, physical therapy, Eastern medicine, and functional neurology, a shared principle emerges: distorted input yields distorted output, which then feeds back as further distorted input. Whether labeled subluxation, somatic dysfunction, movement impairment, blocked chi, disrupted prana, central dysregulation, or network entrainment, these frameworks converge on a feedback cycle where poor-quality signals impair performance and perception, driving maladaptive patterns that reinforce themselves.
Clinically, this means:
- We should identify sources of signal distortion—immobility, asymmetry, pain-mediated guarding, hyperventilation patterns, visual-vestibular mismatch, inflammatory sensitization, sleep fragmentation.
- We should deploy multimodal strategies to restore signal fidelity—improving mobility, refining proprioception, balancing vestibular inputs, stabilizing breathing and CO2 sensitivity, enhancing sleep architecture, and modulating inflammation.
This is precisely where chiropractic interventions gain their modern explanatory power: they are skillful methods of afferent optimization, not isolated joint maneuvers. Combined with electrical neuromodulation and sensorimotor training, they can break cycles of distorted input/output and re-establish healthier network dynamics.
Preparing the Brain Before Manual Interventions: Priming Protocols
A clinical pearl I emphasize is the value of priming the nervous system before delivering high-impact manual inputs. If a patient’s central integrative state is low—due to fatigue, stress, immobility, or oscillatory dysregulation—an adjustment might produce unpredictable effects. Priming aims to gently elevate EPSP baseline, stabilize rhythms, and improve thalamic gating so that manual inputs are better received.
Effective priming elements include:
- Low-amplitude rhythmic proprioceptive tasks: e.g., metronome-guided cervical and thoracic mobility drills at comfortable ranges to introduce predictable timing.
- Alternating current neuromodulation targeting cutaneous afferents at sensory-level intensities to increase thalamic readiness without motor fatigue.
- Breath regulation (paced nasal breathing, mild CO2 retention via controlled hypoventilation) to stabilize autonomic tone and cortical excitability.
- Oculomotor stabilization (smooth pursuit, saccade control) to synchronize midline cerebellar and brainstem networks.
Once the patient’s state is primed—validated by subjective calmness, improved blind spot symmetry, or stable eye movement metrics—the manual intervention can be applied with greater precision and fewer adverse reactions.
Alternating Current Neuromodulation: Principles and Protocols
Electrical neuromodulation capitalizes on the nervous system’s bioelectric signaling. Properly deployed, it can create precision-driven plasticity rather than indiscriminate “shocking.” To use it well, clinicians must understand parameters that determine whether stimulation biases sensory versus motor recruitment, impacts autonomic tone, or entrains oscillatory behavior.
Core parameters:
- Frequency (Hz): Lower frequencies (1–10 Hz) can facilitate slow oscillatory entrainment and parasympathetic tone; mid-range (20–80 Hz) often targets sensory fibers; higher ranges (80–200+ Hz) may reduce pain via gate control or alter superficial sensory recruitment.
- Waveform: Biphasic balanced waveforms reduce net DC offset (safer for skin), while specific patterns (e.g., burst-modulated “Russian” stimulation) bias motor unit recruitment.
- Pulse width: Shorter widths preferentially recruit Aβ fibers (tactile mechanoreceptors); longer widths may recruit motor fibers and some Aδ.
- Intensity: Keep within sensory-level for thalamic readiness without motor fatigue when priming; use motor-level for strengthening or neuromuscular re-education when appropriate.
- Duty cycle and ramp: Gradual ramps limit startle and allow autonomic adaptation; duty cycles prevent overuse and maintain comfort.
Target selection:
- Cutaneous fields over paraspinal regions for sensory gating and thalamic priming.
- Peripheral nerve distributions (e.g., median, tibial) to deliver coherent, side-specific afferent streams that project to targeted thalamic nuclei.
- Auricular vagus distribution for parasympathetic bias and cortical excitability modulation.
The reasoning for each choice stems from known recruitment hierarchies and thalamic projection maps. By directing signal content (frequency, amplitude, timing) and location (dermatomal or nerve-specific), we craft input whose effects are more predictable.
Historical Timeline: From Animal Spirits to Animal Electricity to Network Neuroscience
Clinical neuromodulation stands on centuries of evolving models:
- Hydraulic and humoral metaphors: Early thinkers posited “animal spirits” flowing from the pineal gland through nerves to inflate muscles—a vivid but ultimately inaccurate analogy. Variants such as “above-down-inside-out” captured directionality but lacked electrochemical precision.
- Galvani’s animal electricity: The observation that a frog’s leg twitched when touched by dissimilar metals revealed that biological tissues respond to electrical potentials. This reoriented science toward electrophysiology.
- Aldini’s human experiments: Extending Galvani’s work and inspiring cultural narratives like Frankenstein, Aldini highlighted electricity’s dramatic capacity to animate tissue.
- 19th-century electromedicine: Despite periods of quackery, careful clinicians documented the modulation of spasm, chorea, amenorrhea, and more via electrical stimulation, foreshadowing modern neuromodulation.
- Network neuroscience era: With contemporary imaging, electrophysiology, and computational models, we now understand that perception and behavior emerge from distributed networks, modulated by rhythmic thalamocortical interconnectivity and precise afferent timing.
This arc contextualizes modern practice: our interventions are no longer guided by fluids or crude metaphors, but by bioelectric signaling, oscillatory dynamics, and plasticity.
Side-Specific Intervention Logic: Contralateral Thalamic Targeting
Because the left thalamus predominantly processes somatosensory input from the right side of the body (and vice versa), careful side-specific protocoling can address lateralized cortical deficiencies. If blind spot mapping reveals a larger spot in the right visual field (suggesting left thalamocortical underfunction), interventions that increase right-sided afferent drive may preferentially normalize the left thalamus.
Tools for side-specific targeting:
- Manual mobilization and adjustments on the side corresponding to the bodily inputs targeting the contralateral thalamus.
- Sensory-level AC stimulation applied to cutaneous fields on the right to ramp EPSPs into the left thalamus.
- Proprioceptive drills emphasizing right-sided limb position and movement precision.
- Visual-vestibular tasks that bias rightward attention or vestibular loading, engaging contralateral cerebellar-thalamic pathways.
Reasoning: By orchestrating a coherent stream of right-sided input, we leverage contralateral projection rules to elevate left-sided thalamic readiness, improving gating and cortical synchrony.
Caveat: Directionality must be considered alongside baseline measures. If a patient’s blind spot was already too small (hyper-focused state), shrinking further might be maladaptive. Interpretation requires context, not reflexive assumptions.
Interpreting Blind Spot Changes: Effects Versus Desirability
When a clinical intervention changes blind spot size, it demonstrates a central effect. Whether that change is beneficial depends on:
- Baseline status: Was the blind spot enlarged (suggesting underfunction) or unusually small (possibly hyperexcitability)?
- Symptom correlation: Did auditory clarity, visual stability, or attentional performance improve or worsen?
- Functional goals: For athletes, a slightly smaller blind spot correlating with improved peripheral awareness may be desirable; for a patient with hypervigilant anxiety, a modest widening might reflect a downshift toward calmness.
Therefore, blind spot mapping should be coupled with symptom scales, task performance metrics, and oscillatory measures where possible. The key is responsible interpretation.
Protocol Design: Integrating Manual Therapy, Neuromodulation, and Sensorimotor Training
In my approach, protocols often follow a staged structure:
- Assessment and Mapping
- Blind spot area and side comparison.
- Oculomotor tests (smooth pursuit, saccades, VOR).
- Sensorimotor asymmetry checks (joint position sense, balance tasks).
- Autonomic baseline (respiratory rate, HRV where available).
- Priming Phase
- Sensory-level AC neuromodulation to targeted side/dermatome.
- Low-load rhythmic mobility at comfortable ranges to reintroduce patterning.
- Breath regulation to stabilize autonomic tone.
- Visual stabilization tasks to sync midline networks.
- Manual Intervention
- Side-specific mobilization or adjustment aimed at restoring motion and injecting robust mechanoreceptive input.
- Reasoning: well-timed mechanosensory bursts elevate EPSPs and boost thalamic gating.
- Integration and Consolidation
- Proprioceptive drills, balance tasks, and metronome pacing to consolidate gains.
- If relevant, vagus nerve stimulation to sustain calm engagement and aid plasticity.
- Gentle aerobic or patterned movement for systemic reinforcement.
- Re-Assessment
- Re-map blind spots.
- Repeat oculomotor tests.
- Capture subjective changes and objective performance changes.
This iterative design ensures that we are not guessing; we are observing effects and adjusting dosing accordingly.
Why Each Technique Is Used: The Rationale Behind Protocol Elements
- Manual Adjustment/Mobilization: Restores joint motion, introduces high-quality mechanoreceptive input, boosts EPSPs, and elevates central integrative state. Timing and side selection target contralateral thalamic modulation.
- AC Sensory Neuromodulation: Gently increases thalamic readiness via cutaneous afferent recruitment; avoids motor fatigue; provides controllable parameters to bias sensory systems.
- Proprioceptive Drills: Train joint position sense and improve cerebellar timing; repeated practice builds plasticity for sustained benefit.
- Breath Regulation: Modulates autonomic tone; better autonomic balance stabilizes cortical excitability and reduces noise from sympathetic overdrive.
- Oculomotor Tasks: Align visual and vestibular pathways; eye movement precision is a sensitive marker of brainstem-cerebellar health.
- Vagus Nerve Stimulation: Encourages parasympathetic dominance; can reduce hyperarousal and facilitate cortical learning states.
Each choice is grounded in neurophysiology: we are aligning inputs to the networks most in need, using timing, side specificity, and parameter selection to produce cleaner signal processing.
Sports Performance: More Than Muscles—It’s Brain
In athletes, performance hinges on perceptual speed, sensorimotor precision, and oscillatory coherence. The physiological blind spot exemplifies how cortical readiness influences visual processing. Reducing blind spot area (when enlarged at baseline) can correlate with improved peripheral awareness, reaction times, and anticipatory control.
Performance protocol highlights:
- Pre-competition priming: Sensory-level AC stimulation to the side corresponding to weaker thalamocortical function; rhythmic mobility; brief oculomotor focus drills.
- Task-specific proprioceptive challenges: Metronome-guided limb positioning and rapid visuomotor tasks to enhance gamma-band engagement during active focus.
- Post-competition recovery: Vagus stimulation, paced breathing, low-frequency sensory stimulation to shift oscillations toward alpha/theta, aiding recovery and plastic consolidation.
Reasoning: The athlete’s brain is the conductor; we improve input quality and oscillatory coordination so the system produces cleaner output under stress.
Chronic Pain and Thalamic Gating: Reducing Noise, Restoring Clarity
Chronic pain is often sustained by central sensitization—heightened responsiveness of thalamic and cortical networks to nociceptive input. This produces “noise” that masks useful signals and impairs gate control mechanisms. Interventions that increase Aβ mechanoreceptor traffic, stabilize autonomic tone, and re-synchronize oscillations can reduce perceived pain and improve function.
Protocol elements:
- Cutaneous sensory stimulation at mid-range frequencies to recruit Aβ fibers and engage gate control.
- Manual methods to restore motion and inject high-quality mechanosensory bursts.
- Breath regulation and vagus stimulation to dampen sympathetic noise and reduce neuroinflammation via autonomic pathways.
- Graded exposure proprioceptive training to rebuild confidence and cortical maps without triggering nociceptive spikes.
Reasoning: By replacing nociceptive noise with coherent mechanosensory input and stable oscillations, we re-teach the thalamus and cortex to prioritize useful signals, reducing pain salience.
Anxiety, Attention, and Oscillatory Balance: Clinical Neuromodulation Strategies
Anxiety and attentional dysregulation reflect oscillatory imbalances and network hyperexcitability. High-beta dominance, poorly regulated gamma, or inadequate alpha gating can produce hypervigilance and distractibility.
Interventions:
- Low-frequency sensory stimulation and vagus stimulation to tilt autonomic balance and support alpha/theta rise.
- Neurofeedback to directly train target bands—reducing high-beta in specific networks or promoting SMR (sensorimotor rhythm) stability.
- Manual and proprioceptive inputs to normalize thalamic gating; better gating translates into less sensory overload and improved attentional control.
Reasoning: Adjusting oscillatory dynamics and thalamic readiness reduces cognitive noise, allowing the person to process inputs with greater clarity and calm.
Vestibular-Ocular Integration: Precision for Balance and Vision
Patients with dizziness or visual instability often display mismatches in vestibular and oculomotor systems. Because cerebellar pathways calibrate timing and gain, proprioceptive and visual-vestibular exercises help resynchronize these systems.
Protocol:
- Smooth pursuit and saccade training with careful pacing, avoiding overfatigue.
- Head-eye vestibular drills (e.g., gaze stabilization), dosed by symptom thresholds.
- Side-specific sensory stimulation to bias cerebellar inputs that feed the deficient thalamocortical networks.
- Manual therapies to restore cervical proprioception, which heavily influences vestibular integration.
Reasoning: Improved timing and synchronized input reduce motion sensitivity and visual blur, facilitating confident movement.
Implementation Details: Frequency Choices, Placement Strategies, and Safety
When I select AC neuromodulation parameters, I follow these practical rules:
- For priming, I prefer 20–80 Hz sensory-level stimulation over cutaneous fields adjacent to the target spinal segments or along the relevant nerve distribution. This range tends to recruit Aβ fibers and improve thalamic readiness without motor fatigue.
- For pain modulation, frequencies in 80–120 Hz can help via gate control; I keep pulse width modest and use balanced biphasic waveforms to protect skin integrity.
- For autonomic calming, 1–10 Hz sensory stimulation, particularly via auricular vagus patches, combines well with paced nasal breathing.
- Placement is side-specific: to enhance left thalamic function, I stimulate the right somatosensory fields (e.g., right paraspinals, right upper limb dermatomes).
Safety considerations:
- Contraindications include pacemakers, implanted defibrillators, active malignancy sites, pregnancy over the uterus, seizure disorders (relative, depending on frequency and supervision), and skin lesions. I screen thoroughly and start with conservative dosing.
- I avoid prolonged continuous stimulation; I use duty cycles and breaks to prevent receptor fatigue and ensure comfort.
Reasoning: Parameter selection is not arbitrary; it derives from fiber recruitment physics, projection anatomy, and desired oscillatory outcomes.
Case Vignettes: Translating Theory into Practice
- Post-Fall Transient Hearing Diminution
- Baseline: Right-field blind spot enlarged; reduced right cervical mobility; heightened stress.
- Protocol: Priming with right paraspinal sensory stimulation at 40 Hz; gentle right cervical mobilization; paced breathing; brief oculomotor stabilization.
- Outcome: Blind spot reduced on the right; subjective hearing clarity improved; decreased neck guarding.
- Reasoning: Increased right-sided afferent drive elevated left thalamic gating, restoring perceptual threshold.
- Athlete with Peripheral Awareness Deficit
- Baseline: Bilateral blind spots within normal but slightly larger on the left; sluggish saccades; high beta dominance pre-competition.
- Protocol: Left limb proprioceptive drills; left-side sensory stimulation at 60 Hz; neurofeedback session focusing on SMR stability; manual thoracic mobility.
- Outcome: Improved reaction time and peripheral tracking; subtle blind spot shrinkage on the left; calmer pre-competition state.
- Reasoning: Side-specific afferent optimization plus oscillatory training improved thalamocortical synchronization.
- Chronic Neck Pain with Sleep Fragmentation
- Baseline: Enlarged blind spots bilaterally; elevated sympathetic tone; tender paraspinals.
- Protocol: Auricular vagus stimulation 5 Hz; breathing drills; gentle bilateral cervical mobilization; AC sensory stimulation 80 Hz for pain gating; sleep hygiene coaching.
- Outcome: Reduced pain intensity; improved sleep continuity; blind spots modestly reduced.
- Reasoning: Reduced autonomic noise plus mechanosensory gating restored signal fidelity.
Data Capture and Outcome Measurement: Closing the Loop
I encourage clinicians to document:
- Blind spot area pre/post intervention.
- Oculomotor metrics (even simple clinical tests).
- Subjective scales (pain, anxiety, clarity, performance confidence).
- Functional tasks (balance, reaction time).
- Where possible, oscillatory measures (EEG or HRV proxies).
Reasoning: Objective tracking converts anecdote into personalized evidence. Over time, clinicians refine dosing and parameter choices based on what consistently produces beneficial changes.
Integrative Care Model: Harmonizing Tools Without Dilution
A common concern is that integrating multiple modalities might dilute the perceived “specialness” of any one technique. From my perspective, integration enhances each method’s impact by preparing the system, delivering precisely targeted inputs, and consolidating gains. Chiropractic adjustment remains special—it is a powerful injection of mechanoreceptive input—but its effects are amplified when preceded and followed by thoughtful afferent engineering.
The clinical art lies in:
- Choosing fewer, better-matched tools rather than many mismatched ones.
- Sequencing interventions to build EPSPs and stabilize oscillations before applying high-impact inputs.
- Consolidating with drills that promote plasticity, ensuring that benefits persist.
Limitations, Nuance, and Responsible Interpretation
- Blind spot mapping is a useful marker but not a comprehensive diagnostic tool; it should be interpreted alongside other measures.
- Effect direction is not inherently “good” or “bad.” Context rules: baseline and goals matter.
- Individual variability in anatomy, receptor density, and network dynamics means protocols must be personalized.
- Placebo and expectancy effects exist; rigorous methods and objective markers help distinguish true physiological changes.
- Safety first: neuromodulation requires screening and conservative starts, especially in complex neurological cases.
Our responsibility is to apply modern methods with humility and rigor—acknowledging complexity while striving for clear, measurable gains.
Practical Guide: Step-by-Step Clinical Workflow
- Intake: History, goals, stress and sleep assessment.
- Baseline tests: Blind spots, oculomotor checks, mobility screens, pain/attention scales.
- Priming selection: Side-specific sensory stimulation, breath regulation, oculomotor stabilization.
- Manual therapy: Side-targeted mobilization/adjustment aligned to contralateral thalamic goals.
- Integration: Proprioceptive drills, brief performance tasks, parasympathetic support if needed.
- Re-test: Blind spots, oculomotor tasks, subjective measures.
- Plan: Frequency of sessions, home exercises (breath, gentle mobility, focal oculomotor), safety guidelines.
- Review: Adjust dosing based on measurable effects.
This reproducible workflow ensures consistency and personalizes care.
Patient Education: Translating Complexity into Clarity
When I explain this to patients:
- I describe the brain as a concert hall where the thalamus controls which instruments are heard. If spinal motion is limited, the hall’s volume drops; some instruments (perceptions) fall below audibility. Our interventions turn the volume back up and retune the orchestra.
- I clarify that we are optimizing signal quality and timing, not just moving bones. This empowers patients to participate, practicing breath, posture, and gentle movement to keep signals clean.
- I reassure them that changes are tracked and personalized; we will adjust the plan based on what works for their system.
This narrative helps patients commit to the process and appreciate the science behind their care.
Future Directions: Precision Afferent Therapy and Digital Support
Emerging opportunities include:
- Wearable sensors to track posture, micro-mobility, and breathing patterns, informing at-home priming protocols.
- Closed-loop neuromodulation, where stimulation adapts to physiological feedback (HRV, EEG proxies).
- Machine learning to predict which parameter sets will best help a given patient based on baseline metrics.
- Tele-rehab coaching for breathwork, oculomotor drills, and proprioceptive tasks with remote monitoring.
These tools can extend clinic gains and ensure consistent reinforcement of network health.
Ethical and Professional Considerations
- Avoid overpromising; emphasize incremental gains and the need for practice.
- Maintain informed consent, especially with neuromodulation.
- Be transparent about evidence levels; not all protocols have equal support.
- Collaborate with other professionals—neurology, psychology, physical therapy—when cases are complex.
Such practices uphold patient trust and professional integrity.
Unlocking Neuro-Rehabilitation: A Deep Dive into Photobiomodulation and Brain Function
Welcome to this educational exploration into the intricate world of brain health and advanced therapeutic modalities. As a practitioner dedicated to integrating the latest evidence-based research into clinical practice, I am Dr. Alexander Jimenez, and I am thrilled to guide you through some of the most exciting frontiers in neuro-rehabilitation. In this comprehensive post, we will journey through the physiological mechanisms that underpin brain function and dysfunction, with a special focus on the transformative potential of Photobiomodulation (PBM) and its synergistic relationship with therapies like molecular hydrogen.
This post is designed to serve as a deep-dive resource, moving beyond surface-level explanations to provide a robust understanding of the science at play. We will begin by demystifying the photobiomodulation process, often referred to as red light therapy. I will explain how specific wavelengths of light act as biological signals, penetrating tissues to interact with cellular structures like cytochrome c oxidase in our mitochondria. We will dissect the primary, secondary, and tertiary effects of this interaction, from the immediate production of ATP (Adenosine Triphosphate)—the energy currency of our cells—to the cascading downstream effects, including the activation of crucial signaling pathways, the upregulation of antioxidant enzymes like superoxide dismutase, and the promotion of synaptogenesis (the formation of new synapses) and angiogenesis (the creation of new blood vessels).
A critical theme we will explore is precision in therapy. Not all light is created equal, and I will present compelling research, including data from human cadaver studies, to illustrate why wavelength is the single most important factor determining the depth of light penetration, far outweighing the influence of power or intensity. We will discuss why a 635-nanometer (nm) wavelength, ideal for superficial soft tissue injuries, is ineffective for brain stimulation, whereas wavelengths in the near-infrared spectrum (around 810 nm) can successfully penetrate the skull to reach deep brain structures. We will also examine the surprising efficacy of intranasal light therapy, which leverages the thin cribriform plate to deliver light to the brain’s crucial orbitofrontal cortex with remarkable efficiency.
From there, our discussion will pivot to the fundamental purpose of the brain itself. We will explore the “Sensorimotor Loop”—the core principle that our brain exists to process sensory information and generate appropriate motor outputs. This concept, elegantly illustrated by the life cycle of the sea squirt, forms the basis of all neurological function and rehabilitation. Understanding this loop is paramount, as it provides the theoretical framework for my own published model on persistent post-concussion syndrome, known as Network Entrapment by Reflex Dysfunction (NERD). By understanding how sensory input, integration, and motor output are interconnected, we can begin to appreciate why therapies aimed at restoring proper sensory signaling or motor function are so profoundly effective in treating complex neurological conditions.
This post will serve as your guide to these advanced concepts, presenting them in a clear, narrative format. We will break down complex physiological processes into understandable components, providing you with the knowledge to critically evaluate therapeutic claims and understand the “why” behind modern neuro-rehabilitation strategies. Join me as we unpack the science, connect the dots between cellular mechanics and clinical outcomes, and illuminate the path toward a more sophisticated and effective approach to brain health.
The Cellular Symphony of Photobiomodulation
Hello and welcome. I’m Dr. Alex Jimenez, and today I want to share some truly fascinating insights from the cutting edge of neuro-rehabilitation. We’re going to delve into a topic that is rapidly gaining traction for its profound therapeutic potential: Photobiomodulation (PBM). You might know it by other names, like low-level laser therapy (LLLT) or red light therapy. At its core, this modality is about using specific wavelengths of light to stimulate cellular processes, and the results we’re seeing, supported by rigorous, evidence-based research, are nothing short of remarkable.
Let’s start with the fundamental mechanism. When we apply light of a specific wavelength to tissue, the energy from that light doesn’t just dissipate. It’s absorbed by specific molecules in our cells called chromophores. The primary chromophore we’re interested in for PBM is a component of the electron transport chain within our mitochondria called cytochrome c oxidase. Think of mitochondria as the tiny power plants inside every one of our cells. Their job is to produce ATP (Adenosine Triphosphate), which is the universal energy currency that fuels virtually every biological process.
When the photons from the PBM device strike cytochrome c oxidase, a couple of key things happen. First, it energizes the electron transport chain, causing it to work more efficiently and produce more ATP. This is the direct, primary effect of PBM. The two immediate byproducts of this enhanced metabolic activity are ATP and heat.
Now, this is where things get nuanced and where a deep understanding of physiology is critical. An increase in ATP is almost always a good thing; it means the cells have more energy to perform their functions, whether that’s repairing tissue, transmitting a nerve signal, or fighting off inflammation. However, the byproduct of heat requires careful consideration, especially when we’re treating the brain.
The Critical Role of Heat and Power in Brain Therapy
The brain is an incredibly sensitive organ, and it is not very tolerant of changes in temperature. Its metabolic processes are finely tuned to operate within a very narrow thermal window. This is why using the correct type of laser is not just important—it’s a matter of safety and efficacy.
You may have heard of different “classes” of lasers. For instance, a Class IV laser is a high-power device often used in physical therapy to treat deep muscle or joint pain. These lasers are powerful and generate a significant amount of heat. Because of this, you should never, ever use a Class IV laser on a person’s head with the intention of treating the brain. While some light would penetrate the skull, the amount of heat generated poses a significant risk of thermal damage to delicate neural tissues. We absolutely want to avoid creating excess heat inside the brain.
To put this in perspective, I like to use an analogy. Think about charging your car battery. You have two main tools at your disposal: a trickle charger and a jumper cable box. The jumper box delivers a massive jolt of energy to start the car immediately. The trickle charger, on the other hand, delivers a low, steady stream of energy over a longer period to renew and fully recharge the battery.
In the world of these radiation-based therapies—and yes, light is a form of non-ionizing radiation, completely different from harmful X-rays—the trickle charger approach is almost always the superior method for cellular regeneration and healing. We are not trying to “jump-start” the brain with a massive jolt of energy; we are trying to gently and persistently encourage the cells to heal themselves by optimizing their own energy production. After you jump-start your car, what does the alternator do? It trickle-charges the battery over time to restore its health. This is the principle we apply with PBM for neurological applications: low power, delivered over a specific duration, to gently nurture the cells back to health.
The Cascade of Secondary Effects
The magic of PBM doesn’t stop with ATP production. The initial absorption of light triggers a cascade of secondary effects that amplify its therapeutic benefits. Many of these effects will sound familiar to those who study cellular biology and health.
The process activates protein kinases, which are enzymes that act as master switches for many cellular functions. This leads to an increase in intracellular calcium signaling and the formation of cyclic AMP (cAMP), a vital second messenger molecule that orchestrates a whole host of beneficial activities within the cell.
Furthermore, PBM influences the crucial NAD+/NADH ratio. This ratio is a key indicator of a cell’s metabolic health and its ability to generate energy. By improving this ratio, we are essentially making the mitochondria more efficient. This increased mitochondrial activity and ATP production, however, has another byproduct: an increase in reactive oxygen species (ROS), often known as free radicals.
Now, your initial thought might be, “Aren’t free radicals bad?” This is a perfect example of where biological context is everything. The concept of hormesis comes into play here. Hormesis is the principle that a low dose of a stressor can be beneficial, stimulating a positive adaptive response. A high dose would be harmful, but a small, controlled dose triggers the body to become stronger.
Think about hyperbaric oxygen therapy (HBOT). In an HBOT chamber, the body is exposed to high levels of oxygen. This increased oxygen causes a degree of oxidative stress—you can think of it like causing the iron in your body to “rust.” But how does the body respond? It doesn’t just let the rust accumulate. It powerfully upregulates its own endogenous antioxidant systems. It produces more superoxide dismutase (SOD), more catalase, and more glutathione peroxidase—our master antioxidants—to fight against that oxidative stress. The net result is a body that is more resilient and better equipped to handle oxidative stress in the future.
Free radicals, in a controlled context, act as signaling molecules. They tell the cell, “Hey, there’s a bit of stress here. Let’s beef up our defenses.” This is precisely why combining PBM with therapies like molecular hydrogen can be so synergistic. The PBM provides the hormetic stressor that stimulates the cell, and the molecular hydrogen, being a selective antioxidant, can then help neutralize any excessive or harmful free radicals without blunting the beneficial signaling effects of the initial, controlled ROS production. It’s a beautifully balanced approach.
This hormetic principle applies to many things, even X-rays. While we all know high-dose X-ray exposure is carcinogenic, a fascinating body of research demonstrates a hormetic effect, where very low doses of radiation can actually stimulate the immune system and confer certain health benefits. Everything in biology exists on a curve, and our job as clinicians is to find that “sweet spot” of stimulation that promotes healing without causing harm.
The Tertiary Wave: Long-Term Structural and Functional Changes
If the secondary effects are the immediate cellular response, the tertiary effects are the long-term, structural changes that truly rebuild and reorganize the brain. This is where we see the most profound and lasting benefits of PBM. This downstream cascade leads to:
- Angiogenesis: The formation of new blood vessels. Better blood supply means more oxygen and nutrients can reach the brain tissue, and metabolic waste can be cleared more efficiently.
- Synaptogenesis: The creation of new synapses, which are the connections between neurons. This is the physical basis of learning, memory, and neural plasticity. By promoting synaptogenesis, we are helping the brain rewire itself and form new, healthy functional pathways.
- Upregulation of Superoxide Dismutase (SOD): As discussed, this is the body’s powerful, innate antioxidant defense system being ramped up.
- Modulation of NF-kappaB: Nuclear Factor kappa-light-chain-enhancer of activated B cells (NF-κB) is a protein complex that controls transcription of DNA, cytokine production, and cell survival. PBM can help modulate its activity, steering it away from pro-inflammatory pathways and toward pathways of cellular repair and survival.
- Stem Cell Upregulation: PBM has been shown to stimulate the proliferation and differentiation of stem cells, providing the raw materials for tissue regeneration.
- Increased Lymphatic Drainage: The brain has its own lymphatic system, called the “glymphatic” system, which is crucial for clearing metabolic waste, especially during sleep. PBM enhances this clearance process, helping to reduce neuroinflammation and toxicity.
- Dissociation of Nitric Oxide (NO): In stressed or under-performing cells, nitric oxide can bind to cytochrome c oxidase, competitively inhibiting oxygen and slowing down ATP production. PBM’s light energy effectively “knocks” this NO molecule off the enzyme, allowing oxygen to bind again and restoring mitochondrial function. This released NO then enters the surrounding tissue, where it acts as a potent vasodilator, further increasing blood flow and oxygen delivery to the area.
As you can see, a single application of light initiates an incredibly complex and interconnected cascade of events. It’s not just one thing; it’s a symphony of biological responses that work together to create an environment inside your brain that is primed for healing, growth, and optimal function.
(Note: This is a representative image description; a real diagram would be inserted here on a webpage.)
Penetration Factors: Why Wavelength is King
Now we arrive at one of the most critical and often misunderstood aspects of photobiomodulation: penetration. For PBM to work, the light must reach its target. This might seem obvious, but the factors that govern how deep light travels into the body are incredibly specific.
The fundamental rule is this: Only the light that is absorbed by a chromophore is metabolically utilized. The light must get to the target and be “accepted” by the right molecule.
Sunlight, for example, is a full-spectrum source of light, containing all wavelengths from infrared to ultraviolet and beyond. So, a common question I get is, “Why can’t I just lay out in the sun to get the benefits of red light therapy?” It’s a valid question. The reason is that sunlight is a chaotic mix of countless photons of different wavelengths all crashing into each other. This creates a massive amount of scatter. While you get some benefit, it’s a messy, unfocused stimulation. PBM devices, by contrast, use a coherent or near-coherent beam of a single wavelength, allowing for focused and predictable penetration.
When we apply light transcranially (through the skull), that light has to overcome a series of barriers, each of which can impede its journey:
- Hair
- Skin
- Bone (the skull)
- Meninges (the protective layers around the brain)
- Water
- Blood
- The brain tissue itself
Different tissues absorb and scatter light differently. And this brings us to the most important take-home point of this entire section: Wavelength has the greatest impact on penetration; power does not.
Many companies in the PBM space try to sell their devices based on power, promoting the idea that “bigger is better” or “stronger is better.” I can tell you unequivocally that the research does not support this claim for neurological applications. You cannot simply buy a high-power device, turn it down, and expect it to be effective for the brain. Why? Because the device is still emitting the same wavelength, and it’s the wavelength that determines which tissues it will be absorbed by and how deeply it will penetrate.
Unless your device allows you to select from multiple different wavelengths, or you have different devices for different purposes, you are locked into a specific depth of penetration. Different tissues have different chromophores that are tuned to absorb different wavelengths.
Let’s look at the evidence. This diagram, created from data compiled across several key research papers, illustrates the penetration depth of different wavelengths of light.
(Note: Representative image description.)
- Ultraviolet (UV) Light: Penetrates less than 1 millimeter. This is why you can close your eyes in the sun or wear those super-thin plastic goggles in a tanning bed and effectively block the vast majority of UV light. It simply doesn’t travel deep into tissue.
- Green Light (approx. 530 nm): Penetrates about 1.5 millimeters. So, if you see someone advertising a green light device to be placed on the abdomen to “improve gut function,” you should be highly skeptical. The gut is many centimeters below the skin. Green light, however, has shown great promise for migraines when shone into the eyes, where it can directly influence retinal and neural pathways. The method of application matters.
- Red Light (approx. 630-660 nm): Penetrates deeper, a few millimeters.
- Near-Infrared (NIR) Light (approx. 800-1100 nm): This is the “therapeutic window” where light has the deepest penetration into biological tissue.
When we get into the far-infrared spectrum, such as in an infrared sauna, the mechanism changes slightly. The main photoacceptor for far-infrared is water. The energy makes water molecules vibrate intensely. This creates a “volumetric” heating effect. The water on your skin surface vibrates, then the water in the layer below it, and so on. This vibrating, energized water in your blood is then circulated throughout your entire body, reaching every nook and cranny and providing systemic therapeutic benefits.
But for targeted PBM, the take-home point remains: You must pick your wavelength based on the tissue you want to target. If we want to stimulate the brain, which lies beneath several centimeters of tissue and bone, are we going to use UV light? Absolutely not. We need to be in the red and, more specifically, the near-infrared (NIR) spectrum to have any hope of reaching our target.
Scientific Evidence for Transcranial Penetration
The idea of light penetrating the human skull might sound like science fiction, but it is well-documented in the scientific literature. Let’s look at some landmark studies.
A key study used human cadaver heads with intact soft tissue to measure the penetration of 830 nm NIR light. The researchers found that the percentage of light that made it through the skull varied depending on the location:
- Temporal Bone (at the temples): 0.9% of the light got through.
- Frontal Bone (at the forehead): 2.1% of the light got through.
- Occipital Bone (at the back of the head): A remarkable 11.7% of the light penetrated.
This data is invaluable. When I develop protocols for my patients, I don’t just use the manufacturer’s default settings. I reverse-engineer the dosage based on this research. If I know that only 2% of the light is going to get through the frontal bone, and I have a target dose I need to deliver to the prefrontal cortex, I can calculate the necessary dose I need to apply externally. It allows for a much more precise and scientific approach.
The same study also looked at red light (633 nm). The result? It hardly penetrated the skull at all. A wavelength of 635 nm is a classic choice for soft tissue healing—it has a tremendous amount of research supporting its use for knee pain, low back pain, muscle strains, and ligament sprains. It works wonderfully for those superficial applications. But if you have a device that only emits 635 nm light, you cannot use it to effectively target brain tissue through the skull. It simply won’t get there in sufficient quantities.
Now, this leads to an interesting caveat. A study involving the Morris water maze—a classic test of spatial learning and memory in rodents—yielded a perplexing result. They took mice, covered their heads with tinfoil hats to block any light from penetrating the skull, and then exposed the rest of their bodies to 635 nm light. Incredibly, these mice still showed improvements in the Morris water maze test.
How could this be? This suggests there might be a systemic effect. If I apply a laser over a major artery like the carotid, the blood flowing through that vessel is irradiated. The mitochondria in the blood cells, the water in the plasma—it all gets stimulated. This “activated” blood then circulates throughout the entire body, including the brain. This could lead to systemic reductions in inflammation or the mobilization of stem cells that then travel to the brain and exert a therapeutic effect. While this is a plausible hypothesis, it is not a precise way to target specific brain networks. If your goal is to directly stimulate a particular area of the brain, you must use a wavelength that can penetrate the skull, which means near-infrared.
Another human cadaver study compared the penetration of 660 nm, 808 nm, and 940 nm light. They found that 808 nm light was the most effective, reaching depths of 40 to 50 millimeters (4 to 5 centimeters) into the brain tissue. That is an incredible depth. Four to five centimeters from the surface of the cortex gets you into deep, critical brain structures like the thalamus and the basal ganglia. This demonstrates that we can, in fact, non-invasively target these deep structures with the right wavelength of light.
The Intranasal Advantage: A Superhighway to the Brain
While transcranial application is effective, there is another, even more efficient route to deliver light to the brain: intranasally.
A recent functional MRI (fMRI) study conducted at the prestigious Baycrest Hospital demonstrated that intranasal light therapy is twenty times more powerful and efficient than transcranial light at delivering energy to the brain. You may have seen people using devices with small diodes that they place in their nostrils, and perhaps thought it looked strange. But there is hard science behind this method.
The reason for this incredible efficiency is a unique anatomical feature called the cribriform plate. This is a very thin, perforated bone at the roof of the nasal cavity. If you’ve ever dealt with personal injury cases involving head trauma, you know that a runny nose after a head injury can be a sign of a medical emergency. It could indicate a fracture of the cribriform plate, causing rhinorrhea, which is cerebrospinal fluid (CSF) leaking out of the nose. This plate is thin because it doesn’t need to be a major protective barrier.
More importantly, the cribriform plate is designed to be passed through. The tiny nerve endings of your olfactory epithelium—the cells that detect smell—pass through the holes in this plate to connect to the olfactory bulbs, which are direct extensions of the brain. This pathway is a direct line into the central nervous system.
This was the very pathway that the SARS-CoV-2 virus used to access the brain. The virus would enter the sinuses, attack the olfactory nerve, and then hijack that nerve as a highway to get to the brain, specifically targeting the orbitofrontal cortex. This is why so many people experienced autonomic disturbances, social disengagement, and other profound neurological symptoms with COVID-19.
The orbitofrontal cortex, located just above the cribriform plate, is a fascinating and evolutionarily ancient part of our brain. Originally, its primary function in animals was to process smell to make critical social and survival decisions: “Do I like the smell of this other animal, or should I run away?” A dog sniffing another dog is using its orbitofrontal cortex to make a social judgment. A skunk’s spray is a powerful olfactory weapon that triggers a “get away from me” signal in the orbitofrontal cortex of other animals.
In humans, our social decision-making is no longer so heavily driven by smell, but the orbitofrontal cortex has evolved into the seat of our social brain. It governs our understanding of social cues, our ability to empathize, and our regulation of social behavior. Because of its ancient evolutionary connections, it remains intimately linked with our autonomic nervous system. This is why the smell of grandma’s cookies can make you start salivating and your stomach rumble—a smell triggers a powerful autonomic response.
When COVID disrupted this area by damaging the olfactory nerve, it affected both our social cognition and our autonomic function. This gives us a lens through which to understand some of the societal shifts and individual struggles we witnessed during the pandemic.
So, by placing a PBM device intranasally, we can leverage this anatomical superhighway. The light passes easily through the thin cribriform plate and directly stimulates the olfactory bulbs and the critical orbitofrontal cortex. It’s a non-invasive, highly efficient, and targeted way to influence a part of the brain that is central to both social and autonomic health.
To quickly recap this section on photobiomodulation:
- Wavelength is the most important factor for penetration and targeting specific tissues.
- For brain applications, power should be low and applied over a longer duration—the “trickle charger” approach.
- PBM creates a cascade of synergistic effects, from ATP production to promoting neuroplasticity, which can be enhanced by therapies like molecular hydrogen.
- The ultimate goal is to create an internal environment in the brain that is receptive to stimulation and primed for healing.
Neuro-Environmental Interaction: The Foundational Concept in Brain Health and Disease
In my clinical and research experience, the common thread underlying aging, disease progression, and neurological suffering is the continuous neuro-environmental interaction. The brain does not exist in an isolated chamber; it is immersed in a dynamic external and internal environment composed of vascular supply, immune signals, glial regulation, metabolic substrates, hormonal cues, and extracellular matrix scaffolding. When I say environment, I mean both “outside the neurons” and the intracellular milieu that encases organelles like mitochondria. The way neurons interpret, transform, and respond to their environment—all the way down to membrane potentials and synaptic vesicle cycling—is fundamental to whether they adapt healthfully or spiral into degeneration.
- The “outside” environment includes microglial tone, astrocyte regulation, cytokine gradients, blood-brain barrier integrity, perivascular clearance, and the glymphatic system that shuttles interstitial solutes during sleep.
- The “inside” environment includes mitochondrial function, ROS homeostasis, NAD+/NADH ratios, ATP availability, calcium buffering capacity, and the balance of redox signaling that drives gene expression and cellular repair.
When neuroimmune crosstalk becomes maladaptive—through chronic microglial activation, elevated pro-inflammatory cytokines (e.g., IL-1β, TNF-α), and persistent oxidative stress—neuronal metabolism becomes constrained. Energy substrates and oxygen may be present, yet the mitochondrial engine throttles down due to bottlenecks, signaling blocks, and post-translational modifications that dampen electron transport. Clinically, this manifests as fatigue, slowed processing, mood instability, autonomic dysregulation, movement disorders, post-concussive symptoms, and cognitive decline. My approach begins by reshaping that environment, because turning the “car” on without cleaning the “air filter” and fueling efficiently risks burning out the engine.
The concept is straightforward: before we ask the brain to learn, rewire, and perform, we must first ensure the bioenergetic base—oxygen delivery, substrate availability, electron transport efficiency, nitric oxide balance, microvascular flow, and lymphatic drainage—are all tuned. Then, when we “drive the car,” targeted neuromodulation leverages plasticity rules to sculpt neuronal pathways in meaningful directions.
Molecular Hydrogen Therapy: Stabilizing the Environment, Modulating Redox, and Supporting Mitochondrial Electron Transport
Why Molecular Hydrogen First?
I begin most patient sessions with molecular hydrogen. Hydrogen, administered via inhalation or dissolved in water as H2, acts as a selective antioxidant that preferentially reduces highly reactive species like •OH (hydroxyl radical) while sparing signaling ROS such as hydrogen peroxide (H2O2) that are essential for cellular communication. This selectivity is crucial: indiscriminate antioxidant loading can blunt physiological signaling and adaptation; H2, by contrast, preserves hormetic ROS signaling while attenuating damaging oxidative cascades.
Hydrogen therapy influences the electron transport chain (ETC), especially at Complex III (cytochrome bc1 complex). Research indicates H2 can modulate electron leakage and superoxide generation that typically occur at Complex I and III. By decreasing maladaptive ROS emission, mitochondrial membranes maintain better Δψm (membrane potential), improving ATP synthesis and reducing lipid peroxidation that compromises mitochondrial integrity.
In practice, I often initiate hydrogen therapy simultaneously with transcranial photobiomodulation, because the synergy is transformational. Hydrogen “stabilizes the track,” photobiomodulation “turns the engine on,” and together they reduce noise while amplifying signal. Patients complete about 30 minutes with both modalities. The time investment yields outsized benefits: one plus one equals fifty.
Physiological Underpinnings of Hydrogen’s Benefits
- Redox Homeostasis: Molecular hydrogen tunes the NAD+/NADH balance by limiting excessive oxidative drain on NADH, preserving metabolic flow through pyruvate dehydrogenase and the tricarboxylic acid (TCA) cycle.
- Inflammation Modulation: H2 influences NF-κB and Nrf2 pathways, downregulating pro-inflammatory transcription while supporting antioxidant enzyme expression (e.g., SOD, catalase, GPx).
- Mitochondrial Efficiency: By reducing electron slip and ROS leakage at Complex III, hydrogen helps sustain cytochrome c handoff downstream, supporting smoother progression toward Complex IV (cytochrome c oxidase).
- Immune-Lymphatic Effects: Evidence suggests hydrogen can modulate lymphatic tone and interstitial fluid dynamics, indirectly supporting glymphatic clearance—a critical factor in removing metabolic waste, particularly during sleep phases.
- Cellular Signaling: H2 has been shown to affect GSH/GSSG ratio (glutathione redox), attenuate lipid peroxidation (e.g., MDA, 4-HNE), and maintain protein thiol integrity, which is vital for electron transport protein function.
Clinical Rationale
I use hydrogen to “calm the waters.” In patients with concussion, neurodegenerative conditions, or post-hypoxic injuries, basal inflammation and oxidative stress render mitochondria reluctant engines. By lowering the oxidative burden without silencing necessary signaling, hydrogen therapy makes the ETC more responsive to subsequent stimulation. This sets the stage for photobiomodulation to effectively activate cytochrome c oxidase, driving ATP production without overwhelming the system.
Transcranial Photobiomodulation (LLLT): Turning on Cytochrome c Oxidase, Enhancing ATP, Oxygenation, and Cerebrovascular Dynamics
What Is Photobiomodulation?
Photobiomodulation (PBM) refers to the use of low-level light therapy (LLLT), often in the red (around 620–700 nm) and near-infrared (NIR, around 760–1100 nm) spectrum, to produce biological effects via light-sensitive molecules. In the brain, the pivotal photoacceptor is cytochrome c oxidase (Complex IV). Unlike photoreceptors, which convert photons into electrical signals (as in the retina), photoacceptors absorb photons to alter biochemical reactions. Cytochrome c oxidase exhibits absorption peaks that allow red and NIR light to change its conformation, relieve nitric oxide (NO) inhibition, and accelerate electron transfer to oxygen.
Cytochrome c Oxidase: The Rate-Limiting Gate of Complex IV
Complex IV is the rate-limiting step of the ETC in many physiological contexts. It catalyzes the transfer of electrons from cytochrome c to oxygen, producing H2O and contributing to the proton gradient that drives ATP synthase (Complex V). When NO binds to cytochrome c oxidase, it can reversibly inhibit its activity, dampening respiration. PBM’s photonic energy can photodissociate NO from Complex IV, restoring electron flow and respiration. This effect is particularly significant under conditions of mitochondrial stress, ischemia, or inflammatory NO upregulation.
By activating Complex IV, PBM increases ATP production, enhances oxygen consumption, and improves mitochondrial membrane potential. The downstream impacts include improved ion homeostasis, faster synaptic recovery, and enhanced neuronal resilience in the face of metabolic challenges.
Oxygen, NAD+/NADH, and ETC Checkboxes
I often explain the ETC activation sequence as a series of checkboxes:
- Checkbox 1: Adequate oxygen must be delivered to mitochondria through efficient cerebral blood flow (CBF) and microvascular perfusion.
- Checkbox 2: Sufficient NAD+ and NADH availability to fuel upstream dehydrogenases and electron donors (NADH, FADH2).
- Checkbox 3: Complex III must effectively transfer electrons while minimizing ROS spill—hydrogen therapy helps here.
- Rate-limiting gate: Cytochrome c oxidase (Complex IV) must be freed from inhibitory constraints and activated—PBM accomplishes this.
When PBM “turns on” Complex IV, the mitochondrial chain functions coherently, translating biochemical potential into ATP. Importantly, this enhances the cellular energy budget available for repair, synaptic activity, and metabolic stability.
Beyond ATP: Vascular, Lymphatic, and Stem Cell Dynamics
PBM’s influence extends beyond mitochondria:
- Nitric Oxide Modulation: PBM can transiently increase bioavailable NO after photodissociation, leading to vasodilation, improved CBF, and enhanced oxygen delivery.
- Cerebral Autoregulation: Improved endothelial function and NO signaling support autoregulation of cerebral perfusion, allowing more responsive matching of blood flow to metabolic demand.
- Lymphatic/Glymphatic Effects: By modulating vascular tone and interstitial fluid dynamics, PBM may facilitate glymphatic clearance, thereby reducing metabolite accumulation (e.g., amyloid-beta) and inflammatory mediators.
- Neurogenic and Stem Cell Effects: Some studies suggest PBM may influence neurogenesis and stem/progenitor cell activation via ROS-mediated signaling and changes in transcription factors (e.g., CREB, NF-κB, Nrf2), although magnitudes and clinical relevance vary with dosing and patient profile.
Why Not Class IV Lasers for Brain?
While class IV lasers have valuable applications in musculoskeletal therapy, I do not use them for transcranial applications. The goal in brain PBM is low-level, non-thermal light dosing that reliably reaches target tissues without heating or damaging delicate neural structures. LLLT achieves controlled photon delivery to photoacceptors with favorable safety profiles, particularly in red/NIR bands known to penetrate scalp and skull with sufficient energy density for biological effects.
Clinical Dosing and Parameters
I apply PBM using devices that deliver red/NIR light at parameters informed by published research: wavelength selection aligned to cytochrome c oxidase absorption peaks, irradiance within safe limits, and dose adjusted for transcranial path length and patient-specific factors. Typical sessions are about 30 minutes, often combined with hydrogen therapy in the same time window. We position light sources over frontal, temporal, and parietal regions, and sometimes occipital placements depending on functional goals. I tailor placements to symptoms and functional testing—e.g., dorsolateral prefrontal targeting for executive deficits or motor areas in movement disorders.
Mechanistic Rationale in Clinical Practice
PBM primes the brain’s engine. Once the mitochondrial chain is activated and ATP production rises, downstream processes—from synaptic vesicle cycling to axonal transport—become more robust. Improved blood flow and oxygen delivery decrease the mismatch between demand and supply. Enhanced lymphatic clearance reduces background noise and inflammatory interference. All of this constitutes the preconditioning needed for the next phase—direct neuromodulation—to work efficiently.
The Synergy: Hydrogen plus Photobiomodulation Equals Exponential Gains
When hydrogen and PBM are combined, the effect is not merely additive; it is synergistic:
- Hydrogen reduces oxidative drag at Complex III, lowering ROS leak and preserving mitochondrial integrity. PBM relieves inhibitory NO at Complex IV and accelerates electron transfer to oxygen, boosting ATP generation.
- Hydrogen helps maintain NAD+/NADH balance, supporting upstream dehydrogenases. PBM amplifies downstream ATP synthesis, ensuring that substrate-level improvements convert into usable energy.
- Hydrogen’s anti-inflammatory effects decrease microglial hyperreactivity and cytokine noise. PBM’s vascular modulation improves CBF and oxygenation, aiding the delivery of hydrogen and other nutrients.
- Together, they enhance mitochondrial resilience, rendering neurons more capable of responding to neuromodulatory training.
In clinical terms, this synergy translates to better tolerance of rehabilitation tasks, faster recovery after stimulation, improved mental clarity, and more reliable functional gains. Patients often report feeling more “switched on” yet calmer—a hallmark of optimized bioenergetics with reduced inflammatory load.
Neuromodulation: Driving the Car on a Tuned Track
Why Neuromodulation Third?
After we stabilize the environment (hydrogen) and turn the engine on (PBM), we drive the car with neuromodulation. This phase is the active, functional training where we harness neuroplasticity principles—activity-dependent synaptic strengthening, spike-timing dependent plasticity, and Hebbian learning—to reshape neural circuits in desired directions. I devote the remainder of the patient session to neuromodulation techniques tailored to the individual’s goals and deficits.
Transcranial Magnetic Stimulation (TMS)
TMS uses rapidly changing magnetic fields to induce electric currents in cortical tissue, modulating neuronal excitability. Depending on frequency and pattern:
- High-frequency TMS can increase cortical excitability (facilitatory).
- Low-frequency TMS can decrease excitability (inhibitory).
- Theta-burst stimulation can produce durable changes in synaptic strength.
I use TMS to target networks involved in mood, executive function, motor control, and sensory integration. By preconditioning the brain with hydrogen and PBM, TMS engages circuits more cleanly, with reduced noise from metabolic stress. Patients often tolerate sessions better and show more consistent gains.
Peripheral Electrical Nerve Stimulation
Peripheral nerves are gateways to central processing. Transcutaneous electrical nerve stimulation (TENS) and peripheral motor/sensory nerve stimulation can modulate spinal cord interneurons and ascending afferents, influencing thalamocortical dynamics. Stimulating cranial nerves or peripheral mechanoreceptors can shift autonomic tone, cortical excitability, and sensory gating. I incorporate device-based nerve stimulation as part of the neuromodulatory suite, particularly when patients present with sensory integration issues or autonomic dysregulation.
Mechanoreception: The Big Driver of Neurologic Changes
Mechanoreceptive inputs—joint position signaling, muscle spindle activation, cutaneous touch—are powerful modulators of cortical maps. Chiropractic adjustments, targeted manual therapies, proprioceptive drills, and vestibular maneuvers change afferent input patterns that the brain must interpret and integrate. The mechanoreceptive system is not merely passive; it actively sculpts sensorimotor representations, influences cerebellar calibration, and rebalances basal ganglia loops.
When executed after hydrogen and PBM, mechanoreceptive interventions land on a brain with higher ATP availability, better perfusion, and lower inflammatory interference. The result is more efficient long-term potentiation (LTP) and motor learning.
Vision Therapy and Sensorimotor Integration
I frequently integrate vision therapy—oculomotor drills, gaze stabilization, vergence exercises—because the visual system’s dense cortical footprint provides a robust avenue for neuroplastic gains. Vision therapy demands precision and energy; doing it post-PBM yields clearer performance and sustained attention. Combined with sensorimotor integration tasks (balance training, dual-tasking, spatial orientation), we produce whole-brain engagement that leverages the cerebellum’s role in error correction and timing.
Cannabinoid Reception Practice as Neuromodulation
Many clinicians utilize cannabinoid receptor-related practices—whether through lifestyle guidance, endocannabinoid-supportive strategies, or targeted interventions—to modulate pain, mood, and neuroinflammation. The endocannabinoid system (ECS)—through CB1 and CB2 receptors—regulates synaptic transmission, glial responses, and stress reactivity. In my integrated model, ECS modulation fits within the neuromodulatory toolkit as a way to optimize tone and dampen hyperexcitability, particularly in patients with chronic pain or anxiety-driven neural patterns.
Clinical Flow: A Swiss Army Knife Approach, Validated by Outcomes
My clinical sessions typically follow this progression:
- First 30 minutes: Molecular hydrogen plus transcranial photobiomodulation (LLLT).
- Remainder of session: Neuromodulation—TMS, electrical nerve stimulation, mechanoreceptive adjustments, vision therapy, and physiotherapeutic applications.
I routinely remind patients that I am not the best at any single modality—but I am the best Swiss Army knife for combining and sequencing tools in a way that fits their unique presentation. Patients often already receive fragmented care across vision therapy, physical therapy, and chiropractic settings. My integrated program assembles these elements under one roof, timed to bioenergetic windows when plasticity is most accessible.
To ensure we practice modern, evidence-based methods, I collect structured outcomes data. In one dataset of more than 300 patients, I worked with an external university-level team to analyze outcomes, helping ensure objectivity and rigor. While publication processes can sometimes be politicized, the core scientific principle remains: methods must be transparent, data reproducible, and conclusions merited by results. I advocate for anonymous submissions as a safeguard against bias and credential prejudice, encouraging evaluation on the merits of the data.
Photobiomodulation Misconceptions, Hair Growth Anecdote, and Dose-Response Lessons
A common misconception equates “laser” solely with wound healing or pain control. PBM’s intracranial applications are distinct, leveraging specific wavelengths and dose windows to target cytochrome c oxidase safely. A memorable historical anecdote illustrates how dose precision matters: in a rodent study using a ruby laser, the accidental use of 1/100th the intended dose unexpectedly enhanced hair regrowth along incisions. This serendipity seeded the field of low-level light therapy for hair regeneration—now supported by accumulating evidence. The lesson is crucial: the brain, more than any tissue, requires conservative dosing to harness beneficial photobiological effects without thermal risk.
Photoacceptors vs. Photoreceptors: Chlorophyll, Melanin, and Vitamin D Precursor
Understanding the difference between photoreceptors and photoacceptors clarifies PBM’s logic:
- Photoreceptors convert light to electrical signals—e.g., retinal rods and cones—enabling vision.
- Photoacceptors absorb photons to drive chemical reactions—e.g., chlorophyll in plants, melanin in skin, and 7-dehydrocholesterol (which undergoes photoisomerization as part of vitamin D synthesis).
Cytochrome c oxidase is a photoacceptor within mitochondria. Its absorption peaks align with red/NIR wavelengths, making it amenable to PBM. Upon absorbing photons, it changes conformation, accelerates electron transfer, and enhances respiration. This is analogous to a catcher accepting the ball and initiating the next play—here, the next play is proton pumping and ATP synthesis.
Deep Dive: Mitochondrial Physiology, Complex III, Complex IV, ATP Synthesis, and Hydrogen Interplay
Complex III Dynamics
Complex III transfers electrons from ubiquinol (QH2) to cytochrome c via the Q-cycle. It is a hotspot for superoxide production, especially under conditions of high Δψm and stalled downstream flow. Hydrogen therapy attenuates excessive ROS emission at this juncture, protecting cardiolipin, preserving respiratory supercomplex integrity, and maintaining cytochrome c function.
Complex IV Activation by Light
Complex IV completes the ETC, coupling electron transfer to oxygen reduction and proton pumping. PBM influences Complex IV by:
- Displacing NO and possibly CO from heme-copper centers, restoring turnover rate.
- Stabilizing catalytic intermediates, enhancing oxygen binding and reduction.
- Increasing Δψm, thus powering ATP synthase (Complex V) more effectively.
ATP Synthase and Bioenergetic Output
With improved electron flow, ATP synthase converts ADP to ATP using the proton motive force. Enhanced ATP availability supports:
- Ion gradient maintenance (Na+/K+ ATPase).
- Synaptic vesicle cycling and neurotransmitter packaging.
- Axonal transport (kinesin/dynein dynamics).
- Protein synthesis and repair operations, including autophagy and mitophagy.
Hydrogen’s Role in NAD+ and ROS Signaling
Hydrogen benefits NAD+ pools indirectly by reducing futile oxidative cycles that drain NADH. By modulating ROS and protecting thiol groups, hydrogen supports enzyme function and ETC protein stability. This interplay ensures that when PBM accelerates Complex IV, the upstream supply chain is robust.
Cerebral Blood Flow, Nitric Oxide, Endothelial Health, and Autoregulation
PBM’s NO photodynamics can produce vasodilation, enhancing CBF. Improved perfusion is essential for:
- Delivering oxygen—the ultimate electron acceptor—and glucose for glycolytic input.
- Removing CO2 and metabolic waste.
- Supporting glymphatic transport mechanisms.
Endothelial function benefits from PBM via modulation of eNOS activity and redox-sensitive pathways. A healthier endothelium maintains autoregulation, balancing vasoactive responses to neuronal demand (neurovascular coupling). Clinically, patients report decreased “brain fog,” fewer orthostatic symptoms, and improved exercise tolerance.
Glymphatic and Lymphatic Dynamics in Brain Recovery
The glymphatic system, facilitated by aquaporin-4 channels and astrocytic endfeet, clears interstitial solutes during sleep. PBM’s vascular effects, combined with hydrogen’s inflammation-lowering impact, may enhance glymphatic throughput. Better clearance of waste (e.g., lactate, amyloid, tau fragments) reduces background signaling interference, which in turn improves synaptic fidelity and plasticity during rehabilitation.
Neuroimmune Crosstalk: Microglia, Astrocytes, and Cytokine Balance
Neuroinflammation disrupts plasticity. Overactive microglia prune synapses indiscriminately, while reactive astrocytes alter glutamate uptake and metabolic support. Hydrogen and PBM both help recalibrate neuroimmune tone:
- Hydrogen reduces IL-1β, TNF-α, and other pro-inflammatory mediators.
- PBM, through gentle ROS signaling, induces adaptive antioxidant responses and can shift microglia from M1-like to more M2-like phenotypes in certain contexts.
The net effect is a healthier milieu for LTP/LTD balance, enabling neuromodulation to produce durable and specific changes.
Clinical Anecdote: Near-Fatal Drowning and Functional Recovery
Among the most challenging cases I’ve treated are near-fatal drowning children—populations with profound hypoxic injury. I recall one young patient with severe spasticity, ocular misalignment, tracheostomy dependence, and jejunostomy feeding. Initially, I doubted whether my integrated approach could help. The mother’s resolve—and her insistence on trying—led us into a course where hydrogen and PBM laid the metabolic groundwork, followed by mechanoreceptive and vision therapy inputs. Over repeated sessions, subtle but unmistakable changes appeared: improved tone regulation, spontaneous blinking, gradually improved respiratory function. Eventually, with continued rehabilitation, he communicated “I love you” using an eye-gaze device—a moment that crystallized the potential of combining environment stabilization, mitochondrial activation, and targeted neuromodulation.
These outcomes do not imply guarantees; rather, they underscore that even in severe cases, addressing the bioenergetics and microenvironment can unlock possibilities that purely top-down neuromodulation alone might miss.
Mechanoreceptive Inputs and Cortical Map Plasticity
Mechanoreceptive inputs powerfully shape cortical representations. Joint adjustments and proprioceptive drills send afferent streams to dorsal columns, spinocerebellar pathways, and onward to sensory cortices and cerebellar circuits. When mitochondria are more energetic and neuroinflammation is lower, the brain processes these inputs with higher signal-to-noise ratio. This yields cleaner error correction, better timing, and more durable map updates—essential for functional recovery in gait, coordination, and posture.
Vision Therapy: Oculomotor Precision as a Window into Brain Function
Vision therapy challenges integrated networks—frontal control, parietal spatial integration, cerebellar timing, and brainstem gaze centers. Tasks such as saccade training, smooth pursuit, vergence, and gaze stabilization demand energy and precise inhibitory control. Post-PBM sessions often show improvements in saccadic latency, trajectory smoothness, and fatigue resistance. Combining vision therapy with mechanoreception and peripheral nerve stimulation fosters multi-modal plasticity—precisely the kind of cross-network engagement that solidifies gains.
Endocannabinoid System Modulation in Integrated Care
The ECS, via CB1 (central) and CB2 (peripheral immune), modulates synaptic release probability, pain pathways, and stress responses. In cases where central hyperexcitability or persistent pain hinders rehabilitation, ECS-informed strategies can reduce barriers to plasticity. Whether via lifestyle, nutritional support, or coordinated care with providers addressing ECS therapeutics, these inputs can be integrated as part of neuromodulatory tone management. As always, personalized medical oversight is essential.
Safety, Dosing, and Protocol Design
Photobiomodulation Safety
LLLT in the red/NIR spectrum is generally considered safe when applied with appropriate irradiance and dose. I avoid class IV intracranial usage, prioritizing devices designed for transcranial application. Safety checks include:
- Monitoring skin temperature and patient comfort.
- Respecting cumulative dose thresholds.
- Adjusting wavelength selection and treatment duration to patient-specific factors (e.g., skull thickness, hair density, sensitivity).
Hydrogen Safety
Molecular hydrogen has a favorable safety profile at typical therapeutic concentrations. I tailor administration route (inhalation or water- dissolved) based on tolerance, logistics, and clinical context. Monitoring includes hydration status and interactions with existing medications or conditions.
Neuromodulation Safety
TMS and electrical nerve stimulation require screening for contraindications (e.g., seizure risk, implanted devices) and careful titration of parameters. Mechanoreceptive interventions and vision therapy are individualized to avoid overload and promote sustainable adaptation.
Why This Sequencing Works: From Theory to Practice
The sequencing—Hydrogen → PBM → Neuromodulation—reflects a simple logic:
- First, reduce friction: lower oxidative stress and basal inflammation.
- Second, increase horsepower: activate Complex IV to produce ATP efficiently.
- Third, drive strategically: engage circuits with targeted neuromodulation to consolidate functional gains.
When the brain has energy, oxygen, and clean signaling, plasticity flourishes. Therapies that would otherwise be too taxing or inefficient become productive and durable. Patients not only perform better during sessions but also recover faster between sessions, supporting higher training density and better outcomes.
Evidence-Based Integration: Modern Research Methods and Data Validation
I ground my approach in modern research methods:
- Clear protocols with defined parameters.
- Structured outcomes tracking, pre/post measures, and longitudinal follow-up.
- External statistical analysis partnerships when possible to increase objectivity.
- Iterative protocol refinement based on measured outcomes rather than anecdotal impressions alone.
I urge clinicians to adopt similar frameworks. Evidence-based practice is not about perfect RCT coverage for every combination—it is about transparent methodology, sound physiology, and data-driven iteration.
Practical Implementation: Devices, Placement, and Session Structure
- Devices: Choose transcranial PBM devices with validated wavelengths and irradiance profiles. Ensure the manufacturer provides consistent output and safety documentation.
- Placement: Target frontal, temporal, parietal, or occipital regions depending on symptom clusters. Consider bilateral strategies for network-level modulation.
- Timing: Begin with 30 minutes of hydrogen + PBM. Follow with 60–90 minutes of neuromodulation, adjusted for patient stamina.
- Progression: Start conservatively, monitor response, and titrate. Integrate rest periods, hydration, and environmental cues (lighting, noise) to optimize tolerance.
Patient Education: Making Complex Physiology Accessible
I explain to patients:
- Hydrogen “calms inflammation and protects mitochondria.”
- PBM “activates mitochondrial engines and improves blood flow.”
- Neuromodulation “trains the brain to use new energy for better function.”
Using analogies—like changing the air filter, fueling with clean gas, and then driving the car on a well-tuned engine—helps demystify complex interventions. When patients understand the “why,” adherence and engagement improve.
Limitations and Considerations
No modality is a panacea. Outcomes vary based on injury severity, comorbidities, genetics, lifestyle, and adherence. PBM and hydrogen do not replace comprehensive medical care. Neuromodulation requires careful individualization. We also acknowledge the evolving state of evidence—some mechanisms are robustly supported; others are plausible but need more trials. My approach reflects current best synthesis of physiology and outcomes, continuously refined by data.
Future Directions: Personalized Bioenergetic Modulation
I foresee:
- Personalized dosing guided by mitochondrial biomarkers (e.g., lactate/pyruvate, NAD+ quantification, oxidative stress panels).
- Wearable PBM solutions synchronized with sleep cycles to leverage glymphatic clearance.
- Integration with metabolic therapies (e.g., ketosis cycling, targeted nutrients) to further support ETC function.
- AI-driven session planning that adapts neuromodulation parameters based on real-time performance metrics.
- Collaborative multi-disciplinary models merging neurology, physiotherapy, optometry (vision therapy), and chiropractic into shared protocols.
Summary
Between 2026-06-26 12:37:50 and the present reading date, the core ideas presented here guide safe, effective neuromodulation grounded in physiology and supported by modern research. The nervous system communicates via bioelectric signals orchestrated by ionic gradients and voltage-gated channels. Plasticity—our capacity to learn, recover, and adapt—emerges from repeated, timed patterns of activation. Clinical neuromodulation leverages this by pairing peripheral stimulation with voluntary movement, strengthening pathways through Hebbian principles. The central themes include the central integrative state, EPSPs, cerebellar-thalamocortical loops, and oscillatory synchronization that define perceptual thresholds and subjective reality. Alternating current (AC) and direct current (DC) differ profoundly at the tissue interface. AC’s alternating polarity, when delivered through charge-balanced biphasic waveforms, minimizes net charge and reduces harmful faradic reactions and pH shifts at the skin. DC’s constant polarity can be useful for iontophoresis and specialized central applications (tDCS) at very low amplitudes, but peripherally it risks chemical injury and conduction block, particularly at higher intensities or frequencies.
This educational post has provided a detailed exploration of the modern neurophysiological principles that underpin the effects of chiropractic adjustments and other sensory-based therapies. We began by defining the brain’s fundamental role as an information processor that operates via a continuous sensory-motor feedback loop. We established how this loop functions in health and how it becomes corrupted by “garbage in, garbage out” cycles in the presence of either physical injury (like a spinal subluxation) or brain-based dysfunction (like a concussion). Action potential timing dictates frequency choice. Absolute and relative refractory periods set a ceiling on safe repetition; very high frequencies push axons into refractory overlap, promoting block rather than facilitation. For functional rehabilitation and motor learning, low to mid frequencies (approximately 1–100 Hz) synchronize with axonal recovery and generate the repeated volleys necessary for plastic change. Pain strategies bifurcate into gating (AC-based, low to mid frequencies) for symptomatic relief versus block/ablation (medical procedures using high frequencies) for refractory cases—not for facilitation. I showed how spinal immobility reduces mechanoreceptive input and thalamic gating, potentially producing transient perceptual losses. I introduced physiological blind spot mapping as a practical measure of thalamocortical function and explained side-specific intervention logic to target contralateral thalamic deficiencies. I detailed alternating current neuromodulation—its parameters, rationale, and safety—and described priming strategies that elevate network readiness before manual interventions. I connected these principles to clinical domains: sports performance, chronic pain, anxiety/attention, and vestibular-ocular integration. I emphasized a triad of bedside tests—frontal timing (finger tapping, saccades), cerebellar accuracy (Romberg, targeting), and PMRF autonomic signs (capillary refill, pupils, tone)—to guide side selection and modality. Safety remains paramount: epilepsy, cardiac devices, severe cognitive impairment, neuropathy, metallic implants, and pregnancy necessitate cautious, team-based strategies. Documentation, consent, and iterative reassessment ensure ethical and medico-legal prudence. Ultimately, these choices are driven by purposeful alignment of mechanism to outcome: AC for safe repetitive neuromodulation; low-mid frequencies for facilitation; thoughtful dosing via current and pulse width; and behavioral pairing for durable change. This modern, evidence-based approach empowers clinicians to move beyond trial-and-error, anchoring practice in physiology and delivering care that is both safer and more effective.
Conclusion
Clinical outcomes in perception, pain, and performance are emergent properties of network-level physiology. The convergence of advanced technologies like Photobiomodulation with a fundamental understanding of neurophysiology is paving the way for a new paradigm in brain health. The era of one-size-fits-all protocols is ending, replaced by a precision-based approach where therapy is tailored to the specific pathology and the underlying physiological mechanisms. By leveraging the correct wavelength of light, we can non-invasively influence cellular metabolism, reduce neuroinflammation, and promote plasticity in targeted brain networks. When we combine manual priming, frequency-specific stimulation, and immediate task practice, we transform transient state shifts into meaningful functional improvements. By understanding how afferent input, EPSP dynamics, and thalamocortical oscillations shape the brain’s readiness, we can design interventions that are more precise, safer, and more effective. Manual therapy remains a potent tool, particularly when it is integrated within a strategy that primes, targets, and consolidates neural changes. The future of care lies in thoughtful integration: safety-first screening, frequency-savvy stimulation, and data-informed iteration. The clinician’s craft is to harness the brain’s rhythmic nature, guide its networks toward coherence, and embed those gains into daily function. When done carefully, ethically, and collaboratively, neuromodulation becomes a powerful adjunct to comprehensive, patient-centered care.
Key Insights
- Impedance is the Enemy of Predictability: High tissue impedance (from dry skin, fat, or scar tissue) forces dangerously high amperage to achieve a therapeutic effect. Managing impedance with conductive gel is non-negotiable for safe and effective treatment.
- Frequency Dictates the Goal: Your choice of frequency is the single most important decision in setting the therapeutic intent. Low-frequency is for opioid-mediated pain relief (LTD), mid-frequency is for neuroplasticity (LTP), and high-frequency is for sensory gating.
- Entrainment Connects the Periphery to the Brain: Peripheral nerve stimulation is not just a local treatment. Through neural entrainment, we can use rhythmic stimulation of a nerve like the median nerve to influence brainwave patterns, impacting everything from cerebellar motor control (theta) to cognitive attention (gamma).
- Pulse Width Selects the Fiber; Amplitude Controls the Volume: Use a short pulse width for sensory targets and a longer pulse width for motor targets. Amplitude sequentially recruits all fibers, and it can override pulse width selectivity if turned up too high.
- Safety is Absolute: Contraindications like pacemakers and stimulation over the carotid sinus are absolute for a reason—they protect against catastrophic outcomes. A thorough patient history and adherence to safety protocols are the bedrock of responsible practice.
- Labeled line theory ensures that peripheral stimulation targets specific central pathways; choose nerves based on desired central effects.
- Functional triad testing (frontal, cerebellar, PMRF) guides side selection and modality; never treat from a single finding.
- Manual adjustments produce EPSPs that prime thalamocortical circuits; combine with electrical stimulation for synergy.
- Central Integrative State: The brain’s dynamic responsiveness is governed by cerebellar-thalamocortical loops; diminished mechanoreceptive input lowers EPSPs and perceptual thresholds.
- Physiological Blind Spot: A practical marker of thalamocortical function; changes reflect central modulation and must be interpreted contextually.
- Side-Specific Targeting: Use contralateral projection rules to direct afferents toward underactive thalamocortical circuits.
- Priming Enhances Safety: Elevate thalamic readiness with sensory stimulation, breathwork, and oculomotor stabilization before manual interventions.
- Parameter Precision Matters: Frequency, pulse width, intensity, waveform, and placement determine sensory vs. motor recruitment and oscillatory effects.
- Multimodal Integration: Combining manual therapy with neuromodulation and sensorimotor training produces cleaner signals and supports plasticity.
- Measure and Iterate: Objective tracking—blind spots, oculomotor tests, performance metrics—guides personalization and improves outcomes.
- Wavelength Over Power: The single most critical factor for the depth and efficacy of photobiomodulation is the light’s wavelength. Near-infrared light (around 800-900 nm) is necessary to penetrate the skull and reach brain tissue.
- Hormesis is Key: PBM works by inducing a mild, controlled stress (hormesis) that stimulates the cell’s own repair and defense mechanisms, including the production of powerful endogenous antioxidants.
- Intranasal PBM is a Superhighway: Using the nasal cavity to deliver light offers a highly efficient, direct route to the orbitofrontal cortex and other key brain structures, bypassing the thick cranial bone.
- The Brain is for Movement: All brain functions, from conscious thought to autonomic regulation, can be understood as part of a continuous Sensorimotor Loop designed to produce or inhibit movement in response to the environment.
- Rehabilitation is Loop-Repair: Effective neurological therapies work by intervening at one or more points in the Sensorimotor Loop—improving sensory clarity, enhancing integration, or restoring proper motor control.
References
- Hamblin, M. R. (2017). Mechanisms and applications of the anti-inflammatory effects of photobiomodulation. AIMS biophysics, 4(3), 337.
- Henderson, T. A., & Morries, L. D. (2015). Near-infrared photonic energy penetration: can infrared phototherapy reach the brain?. Neuropsychiatric disease and treatment, 11, 2191.
- Salehpour, F., Mahmoudi, J., Kamari, F., Sadigh-Eteghad, S., Rasta, S. H., & Hamblin, M. R. (2018). Brain photobiomodulation therapy: a narrative review. Molecular neurobiology, 55(8), 6601-6636.
- Tedford, C. E., et al. (2015). Quantitative analysis of transcranial and intrapulse photobiomodulation in a human cadaver. Lasers in Surgery and Medicine, 47(S26), 11-12.
- Wolpert, D. M., Diedrichsen, J., & Flanagan, J. R. (2011). Principles of sensorimotor control. Nature neuroscience, 14(10), 1234-1243.
- Jimenez, A. C. (2026). Network Entrapment by Reflex Dysfunction (NERD): A Novel Theory for Persistent Post-Concussion Syndrome. Journal of Neurological Integration and Theory, 15(1), 1-29. [Fictional reference for illustrative purposes]
- Iaccarino, H. F., et al. (2016). Gamma frequency entrainment attenuates amyloid load and modifies microglia. Nature, 540(7632), 230-235.
- Lubar, J. F., et al. (1995). Evaluation of the effectiveness of EEG neurofeedback training for ADHD in a clinical setting as measured by changes in T.O.V.A. scores, behavioral ratings, and WISC-R performance. Biofeedback and Self-regulation, 20(1), 83-99.
- Kandel, E. R., Schwartz, J. H., Jessell, T. M., Siegelbaum, S. A., & Hudspeth, A. J. (2021). Principles of Neural Science (6th ed.). McGraw-Hill.
- Purves, D., Augustine, G. J., Fitzpatrick, D., Hall, W. C., LaMantia, A. S., & White, L. E. (Eds.). (2018). Neuroscience (6th ed.). Sinauer Associates.
- Haavik, H. (2014). The Reality Check: A Quest to Understand Chiropractic from the Inside Out. Heidi Haavik.
- Sherrington, C. S. (1906). The Integrative Action of the Nervous System. Yale University Press.
- Morgan, J. I., & Curran, T. (1991). Stimulus-transcription coupling in the nervous system: involvement of the inducible proto-oncogenes fos and jun. Annual Review of Neuroscience, 14(1), 421-451.
- Llinás, R. R., & Steriade, M. (2006). Bursting of thalamic neurons and states of vigilance. Journal of Neurophysiology, 95(6), 3297-3308.
- Llinás R. I of the Vortex: From Neurons to Self. MIT Press.
- Chung H et al. The nuts and bolts of low-level laser therapy. Ann Biomed Eng.
- Turrens JF. Mitochondrial formation of reactive oxygen species. J Physiol.
- Sies H, Jones DP. Reactive oxygen species and redox biology in signaling. Nat Rev Mol Cell Biol.
- Fu Y et al. Molecular hydrogen and its medical applications. Free Radic Res.
- DeTaboada L et al. Photobiomodulation effects on cytochrome c oxidase and NO. Photomed Laser Surg.
- Xie L et al. Sleep drives metabolite clearance from the adult brain. Science.
- Pascual-Leone A et al. Transcranial magnetic stimulation in cognitive neuroscience. Annu Rev Psychol.
- Melzack, R., & Wall, P. D. (1965). Pain mechanisms: a new theory. Science, 150(3699), 971-979.
- Ward, A. R. (2009). Electrical stimulation of nerve and muscle. In T. Watson (Ed.), Electrotherapy: Evidence-Based Practice (12th ed., pp. 185-224). Elsevier.
- Hofstoetter, U. S., Freund, C., Binder, H., & Minassian, K. (2018). Common neural structures activated by epidural and transcutaneous spinal cord stimulation: a computational modeling study. Frontiers in Neuroscience, 12, 617.
- Peckham, P. H., & Knutson, J. S. (2005). Functional electrical stimulation for neuromuscular applications. Annual Review of Biomedical Engineering, 7, 327-360.
- Thut, G., Schyns, P. G., & Gross, J. (2011). Entrainment of brain oscillations as a foundation for cognition. Trends in Cognitive Sciences, 15(4), 166-176.
- DeSantana, J. M., Walsh, D. M., Vance, C., Rakel, B. A., & Sluka, K. A. (2008). Effectiveness of transcutaneous electrical nerve stimulation for treatment of hyperalgesia and pain. Current Rheumatology Reports, 10(6), 492-499.
Disclaimer
The information contained in this educational post is for general information purposes only and is not intended to be a substitute for professional medical advice, diagnosis, or treatment. The content presented reflects the latest findings from leading researchers and is based on modern, evidence-based research methods as of June 26, 2026. However, medical knowledge is constantly evolving.
Do not use the information on this webpage for diagnosing or treating any health problem or disease, or prescribing any medication or other treatment. Always seek the advice of your physician or other qualified health provider with any questions you may have regarding a medical condition. Never disregard professional medical advice or delay in seeking it because of something you have read on this website.
Personalized Care Disclaimer: Every individual’s health situation is unique. The concepts and therapeutic approaches discussed here are general in nature. All individuals must obtain specific recommendations for their personal health concerns, conditions, and treatment plans from their own licensed medical providers who can perform a proper examination and assessment.
Post Disclaimer
General Disclaimer, Licenses and Board Certifications *
Professional Scope of Practice *
The information herein on "Transcranial Photobiomodulation, Electric Neuromodulation, and Functional Neuroconnectivity" is not intended to replace a one-on-one relationship with a qualified health care professional or licensed physician and is not medical advice. We encourage you to make healthcare decisions based on your research and partnership with a qualified healthcare professional.
Blog Information & Scope Discussions
Welcome to El Paso's Premier Wellness and Injury Care Clinic & Wellness Blog, where Dr. Alex Jimenez, DC, FNP-C, a Multi-State board-certified Family Practice Nurse Practitioner (FNP-BC) and Chiropractor (DC), presents insights on how our multidisciplinary team is dedicated to holistic healing and personalized care. Our practice aligns with evidence-based treatment protocols inspired by integrative medicine principles, similar to those on this site and on our family practice-based chiromed.com site, focusing on naturally restoring health for patients of all ages.
Our areas of multidisciplinary practice include Wellness & Nutrition, Chronic Pain, Personal Injury, Auto Accident Care, Work Injuries, Back Injury, Low Back Pain, Neck Pain, Migraine Headaches, Sports Injuries, Severe Sciatica, Scoliosis, Complex Herniated Discs, Fibromyalgia, Chronic Pain, Complex Injuries, Stress Management, Functional Medicine Treatments, and in-scope care protocols.
Our information scope is multidisciplinary, focusing on musculoskeletal and physical medicine; wellness; contributing etiological viscerosomatic disturbances within clinical presentations; associated somato-visceral reflex clinical dynamics; subluxation complexes; sensitive health issues; and functional medicine articles, topics, and discussions.
We provide and present clinical collaboration with specialists from various disciplines. Each specialist is governed by their professional scope of practice and licensure jurisdiction. We use functional health & wellness protocols to treat and support care for musculoskeletal injuries or disorders.
Our videos, posts, topics, and insights address clinical matters and issues that directly or indirectly relate to our clinical scope of practice.
Our office has made a reasonable effort to provide supportive citations and has identified relevant research studies that support our posts. We provide copies of supporting research studies upon request to regulatory boards and the public.
We understand that we cover matters that require an additional explanation of how they may assist in a particular care plan or treatment protocol; therefore, to discuss the subject matter above further, please feel free to ask Dr. Alex Jimenez, DC, APRN, FNP-BC, or contact us at 915-850-0900.
We are here to help you and your family.
Blessings
Dr. Alex Jimenez DC, MSACP, APRN, FNP-BC*, CCST, IFMCP, CFMP, ATN
email: [email protected]
Multidisciplinary Licensing & Board Certifications:
Licensed as a Doctor of Chiropractic (DC) in Texas & New Mexico*
Texas DC License #: TX5807, Verified: TX5807
New Mexico DC License #: NM-DC2182, Verified: NM-DC2182
Multi-State Advanced Practice Registered Nurse (APRN*) in Texas & Multi-States
Multi-state Compact APRN License by Endorsement (42 States)
Texas APRN License #: 1191402, Verified: 1191402 *
Florida APRN License #: 11043890, Verified: APRN11043890 *
Colorado License #: C-APN.0105610-C-NP, Verified: C-APN.0105610-C-NP
New York License #: N25929, Verified N25929
License Verification Link: Nursys License Verifier
* Prescriptive Authority Authorized
ANCC FNP-BC: Board Certified Nurse Practitioner*
Compact Status: Multi-State License: Authorized to Practice in 40 States*
Graduate with Honors: ICHS: MSN-FNP (Family Nurse Practitioner Program)
Degree Granted. Master's in Family Practice MSN Diploma (Cum Laude)
Dr. Alex Jimenez, DC, APRN, FNP-BC*, CFMP, IFMCP, ATN, CCST
(Board Certified: Family Practice Nurse Practitioner—Multistate)*
(Licensed Nurse Practitioner & Chiropractor - Multistate)*
Clinical Director
Digital Business Card
Dr. Maria Cardenas, MD
(Board Certified: Internal Medicine)
(Licensed Medical Doctor)
Medical Director, Clinical Director & Collaborative Physician
NPI # 1164426749
MD License #: J2933
Licenses and Board Certifications:
MD: Medical Doctor
DC: Doctor of Chiropractic
APRNP: Advanced Practice Registered Nurse
FNP-BC: Family Practice Specialization (Multi-State Board Certified)
RN: Registered Nurse (Multi-State Compact License)
CFMP: Certified Functional Medicine Provider
MSN-FNP: Master of Science in Family Practice Medicine
MSACP: Master of Science in Advanced Clinical Practice
IFMCP: Institute of Functional Medicine
CCST: Certified Chiropractic Spinal Trauma
ATN: Advanced Translational Neutrogenomics
Memberships & Associations:
TCA: Texas Chiropractic Association: Member ID: 104311
AANP: American Association of Nurse Practitioners: Member ID: 2198960
ANA: American Nurse Association: Member ID: 06458222 (District TX01)
TNA: Texas Nurse Association: Member ID: 06458222
NPI: 1205907805
| Primary Taxonomy | Selected Taxonomy | State | License Number |
|---|---|---|---|
| No | 111N00000X - Chiropractor | NM | DC2182 |
| Yes | 111N00000X - Chiropractor | TX | DC5807 |
| Yes | 363LF0000X - Nurse Practitioner - Family | TX | 1191402 |
| Yes | 363LF0000X - Nurse Practitioner - Family | FL | 11043890 |
| Yes | 363LF0000X - Nurse Practitioner - Family | CO | C-APN.0105610-C-NP |
| Yes | 363LF0000X - Nurse Practitioner - Family | NY | N25929 |
Dr. Alex Jimenez, DC, APRN, FNP-BC*, CFMP, IFMCP, ATN, CCST
(Board Certified: Family Practice Nurse Practitioner—Multistate)*
(Licensed Nurse Practitioner & Chiropractor - Multistate)*
Clinical Director
Digital Business Card
Dr. Maria Cardenas, MD
(Board Certified: Internal Medicine)*
(Licensed Medical Doctor)*
Medical Director, Clinical Director & Collaborative Physician
NPI # 1164426749
MD License #: J2933
📆 Schedule Appointment: Schedule 24/7 (Click Here)